The Origins of the British: The New Prehistory of Britain (53 page)
Read The Origins of the British: The New Prehistory of Britain Online
Authors: Oppenheimer

One possible reason for cultural discrepancies between ‘Saxon homelands’ and Anglo-Saxon England is in the historian’s choice of homeland; Lower Saxony is possibly the wrong place to make such comparisons, and the base of the Cimbrian Peninsula might be more appropriate. However, in the passage quoted above, Hamerow is actually referring to the latter (i.e. the Elbe/Weser region). Another cultural domain, with less self-conscious ethnic marking, is animal husbandry and land use – which, she argues, do ‘suggest a substantial degree of continuity’.
10
Before moving on to the genetic story, perhaps I might allow myself an observer’s comment on archaeological continuity and change in Dark Ages eastern Britain. The one thing on which all archaeologists seem to be agreed is that there were profound changes in English cultural styles, in the broadest sense, over the fifth to seventh centuries
AD
. It would be easier to accept that these could occur without complete replacement if there were pre-existing cultural relationships between eastern Britain and Scandinavia/north-west Europe. The possible elements of these relationships – trade, a common language, prior alliances with high-status marriages – would all contribute to the speed of acculturation and even facilitate a passive acceptance of inward
migration and domination. As discussed earlier, archaeological evidence for such relationships goes back to the Neolithic. It was indeed the main complaint of Gildas in the west that certain ‘tyrants’ actually invited the ‘Saxon’ wolves. Germanic continuity in England would also explain the numerous linguistic anomalies I have been discussing throughout this book, which could be summed up thus: a lack of celtic-language continuity in England.
Two parts of England in particular seem to have been less troubled by the Saxon Advent, and even seemed to have preserved some political continuity, while being ultimately as Germanic, culturally and linguistically, as the rest of Britain: these were Lindsey and Wessex. Lindsey preserved its Roman fort at Lincoln and its Roman name. Roman artefacts persisted until a late date in conjunction with very early Saxon materials in Roman forts at Horcastle and Caistor-on-the-Wolds.
11
Was this evidence just of Germanic mercenaries, or of pre-existing Germanic settlements? Wessex has only patchy archaeological evidence of early Germanic settlers, with complete absence in south Hampshire. John Myres, a traditional invasionist, suggested that an early Wessex king, Cerdic, was the head of a partly British noble family with blood ties to existing Saxon or Jute settlers who had been entrusted with the defence of Wessex in the last days of British sub-Roman authority.
12
Yet, by the time their first literature appears, with his descendant King Ælfred, Wessex had its own local Germanic dialect, with no trace of any celtic lexical substratum.
In pursuing the theme of invasions from north-west Europe, it would be more logical for me to go straight to the last two waves, of Vikings and Normans, and include those with Anglo-Saxons
before looking at the genetic evidence together. They all happened within six hundred years of one another and were, after all, only part of a series of such events which have occurred since the Neolithic. But I shall break here because there are several subjective reasons why the Anglo-Saxon ‘invasions’ should be dealt with on their own. With its dramatic cultural-linguistic schism from Roman Britain, the Anglo-Saxon Advent has led historians and geneticists to cross a speculative chasm on a bridge of flimsy evidence. The resulting extreme view of the slaughter of ‘Celts’ has fulfilled and reinforced Gildas’ doomladen prophecy and his anti-Saxon agenda. This hypothetical holocaust has some psychological significance for the English. The subsequent invasions of the Vikings and Normans, and the occasional subsequent bloody intrusions into remaining non-English-speaking regions by their political descendants, have had perhaps an even greater impact in terms of alienation for the other surviving tribes of the British Isles.
The genetic picture has some colourful history to it as well, for it has been known for almost as long as genetic studies have been possible that there is a genetic fault line between the English and the Welsh that roughly follows the line of Offa’s Dyke. The big questions, as I have tried to stress in this book, are not about whom, but about when, and over what periods, this fault line was created.
Blood groups were one of the first available genetic marking systems that physical anthropologists used to try to categorize the human ‘races’. As it happens, their lack of of variety and their presence, at different proportions, in all populations make
them generally useless for looking in a detailed way at differences and migrations. One of the earliest studies, conducted over eighty years ago, foundered in a morass of absurd relationships, such as Russians linked with Madagascans. Blood groups are still used in population genetic studies, but only as an adjunct to many other marker systems.
13
The genetic difference between England and Wales, however, is so clear that it can be detected by many crude markers, including blood groups. Arthur Mourant, one of the founders of modern physical anthropology, wrote extensively on the biological anthropology of blood groups in the middle of the last century; his autobiography (1995) was aptly entitled
Blood and Stones
. He was not the first to note the higher rates of blood group O, as opposed to A, in Basques, Bretons, Welsh, Irish and Scots, but in 1952, with Welsh colleague Morgan Watkin, he made a reasonable crack at suggesting why:
[T]here appear to us reasonable grounds for the belief that, prior to the advent of Celtic-speaking immigrants, the British Isles were inhabited by a people whose domain had at one time extended over a considerable part of Europe and North Africa but who under ever increasing pressure from the east had been driven from their homelands. Some, no doubt, found refuge in the more isolated mountain regions, but the remainder were gradually driven westwards and finally came to occupy a limited area near the Atlantic seaboard of Europe.
14
Watkin later added:
One wonders, therefore, whether a large part of Britain’s very early population did not arrive by the western sea routes and whether Celtic speech was acquired from later invaders …
15
If we put the language interpretation to one side for the moment, we could say, so far so good: these very general statements, without specifying dates or putting numbers to differences between regions, could be consistent with some of the mitochondrial and Y-chromosome results discussed in this book. However, this blood group evidence has been cited more recently and taken very much further than is warranted. I should like to look at this type of approach in some detail, since it still has lessons for the genetic debate on the ‘Anglo-Saxon replacement’ hypothesis today.
The German linguist Wolfgang Viereck of the University of Bamberg (where blood groups were first discovered) asks: ‘Is there a connection between blood group and membership in a tribe or race and between blood group and language, even between dialectal differences within individual languages?’
16
He then proceeds to build a castle of cards based on correlating minor variations in English dialect with subtle variations in blood group O and A frequencies in Britain. While the dialect variations are interesting, the genetic–linguistic correlations will inevitably end on the heap. However, the use of genetics in this example allows us to look at the use and misuse of genetics and interpretations of how gene frequencies in England could have come to be measurably distinct from those of Wales, Scotland and Ireland.
The basis of Viereck’s geographical comparisons is a map of relative frequencies of blood groups A and O in Britain (
Figure 11.1
). This frequency map is derived from a standard text on blood groups in the United Kingdom.
17
However, the map is not only confusing, it is also a statistically misleading presentation. In all regions except southern England, Norfolk, Lincoln and Nottinghamshire and the two south-western peninsular regions of Britain, blood group O is shown as more common than blood group A. The percentages given in the figure reflect the degree to which one of these two groups exceeds the other, and are intended to indicate the degree of genetic intrusion by A. What they actually do is exaggerate the differences as a result of misunderstanding the genetics of blood groups.
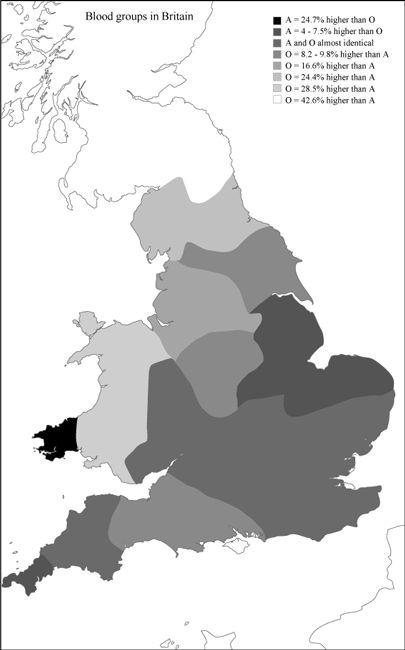
Figure 11.1
Viereck’s map of British blood groups. Drawn from blood bank data, this map purports to show massive ‘younger Germanic blood group A’ invasion into ‘older Celtic blood group O’ in eastern England. The fallacies are: exaggeration of ‘A’ gene frequency, false racial labelling, and no allowance for similar previous colonization history of England and the nearby Continent.
The impression this diagram gives is that in England, blood group A – which in Viereck’s words is ‘said to be the younger Germanic blood group’
18
– has partially replaced blood group O, which he regards as older and ‘Celtic’. This story is wrong, for a number of reasons. For a start, you cannot date blood groups. Group A cannot be specifically ‘Germanic’, nor ‘O’ specifically ‘Celtic’, whatever those designations mean. The gene coding for blood group A is found throughout the world and is no more common in Germany (25–30% allele, or gene frequency) than any other region in mainland Western Europe. The highest frequencies of the A gene type are in fact found among the Saami (35–40%) and to a lesser extent among Scandinavians (30–35%). The group O gene, conversely, is the most common blood group gene type in the world, and outnumbers group A genes nearly everywhere. In Western Europe, the group O gene dominates at 60–70%, and at slightly higher frequencies of 70–80% in the Atlantic fringe. The O gene type in fact greatly outnumbers the A throughout Britain, including England.
19
We might ask how Viereck’s map could get it so wrong and overstate blood group A so much in England (
Figure 11.1
). The answer is simply that the map is based on
blood group
frequencies, not on
blood group gene
frequencies. Our expression of blood groups in the blood typing test depends on the genes we get – one from each parent (our
genotype
). Blood group A is
dominant
, which means that the result of our blood test (our
phenotype
) is group A, whether we get one or two A genes from our parents. Group O is
recessive
, which means that we must receive both our parental genes as O to have an O blood group phenotype. If one parent gives us an O and the other gives us an A, we will test as A. Therefore analyses of blood group frequency will automatically overstate A gene frequencies if this very basic fact is overlooked. Since B, although dominant as well, is quite uncommon in Western Europe, the division of simple blood group frequencies between A and O is a two-horse race, in which the dominant A has the advantage. Small increases in the A gene frequency will be reflected in dramatic relative falls in the O group frequency.
So, where Viereck’s map misleads is in presenting blood group data (phenotypes) rather than gene frequencies (genotypes). Even in England, where blood groups A and O appear to be neck and neck, the group O gene actually still outnumbers A in frequency by up to 2:1. Although the differences in gene frequency probably do represent relative differences in migration history, for the reasons given above they overstate the case and cannot be taken as evidence of recent massive replacement of O by A. This argument would still be a statistical fallacy, even if the A gene were specifically Germanic or the most common gene in Germany were A – neither of which is true.
My main reason for this digression is to show that we can use the simple blood group data to illustrate a major flaw in efforts to determine the size of the Anglo-Saxon or any other Germanic migration or replacement genetically. In a nutshell, the hypothetical Germanic ‘source regions’ were already made up of mixed populations, derived from at least two remote Ice Age refuges shared with the ancestors of people destined for England. This problem also affects analysis of the mtDNA and Y-chromosome data. Let us say (for argument’s sake only) that high rates of the O allele really did represent early post-LGM migrations into Northern Europe from the Basque refuge, and that higher rates of the blood group A allele had arrived in north-west Europe by sailing up the Danube from the Balkan refuge, or at least from Eastern Europe. In this situation, what mix of these two genes would we expect to find in, say, north-west Germany or Frisia?