The Cerebellum: Brain for an Implicit Self (4 page)
Read The Cerebellum: Brain for an Implicit Self Online
Authors: Masao Ito
Tags: #Science, #Life Sciences, #Medical, #Biology, #Neurology, #Neuroscience

We traditionally consider voluntary movements as a much higher order of movements than reflexes in the sense that they are controlled by “free will” and can be performed both automatically and at the level of conscious awareness, whereas reflexes are driven by peripheral stimuli and executed solely by automatic means. However, as our understanding advances for neuronal mechanisms underlying voluntary movements, distinctions between such movements and reflexes become blurred because many of the same neuronal circuits are employed for both types of movement (
Prochazka et al., 2000
;
Hultborn, 2001
). Practically speaking, however, we may still distinguish voluntary movements as initiated from the cerebral cortex, whereas reflexes operate largely within the spinal cord and brainstem. Typically, two cortical areas, the primary motor cortex and the frontal eye field, are involved in voluntary movements of the limbs and eyes, respectively (
Chapters 13
and
14
). In the systems control parlance emphasized in this volume, reflexes and voluntary movements may share neuronal circuits for their controller and controlled object structures, but they are separated from each other by the nature of the instruction signals that drive the controller. Instructions for reflexes arise from periphery, whereas voluntary movements are driven by “top down” instruction signals generated in higher centers of the cerebral cortex, including but not limited to the supplementary motor cortex and the anterior cingulate gyrus (see
Chapter 13
, “
Voluntary Motor Control
”).
An interesting idea has been put forward to suggest that a central instruction causes a voluntary movement by an imitation or replacement of the peripheral stimulus that induces a reflex (the imitation hypothesis; Berkinblit et al., 1986). For instance, the CNS can voluntarily elicit a saccadic eye movement by means of the
imitation of the visual signals that could elicit the saccadic movement reflexively. In this sense, the central instruction may imply an “afference copy” of the peripheral stimulus. Such a capability of imitating a peripheral stimulus might emerge during evolution to develop a neuronal mechanism of voluntary motor control. Neuronal mehanisms underlying the postulated capability of imitation are unknown, but one may suppose that a group of neurons memorize those signals of peripheral stimuli that evoke a motor behavior reflexively and reproduce the same signals whenever a similar motor behavior is to be generated voluntarily. Here, one may recall the “mirror” neurons, which are present in certain cerebral cortical areas and are activated during both observed and performed hand actions, as discussed below (
Section 8
and also in
Chapter 16
, “
Motor Actions and Tool Use
,”
Section 5
). These neurons appear to memorize perceptive signals representing certain successful motor actions performed by another individual and reproduce them as central instructions for their own body’s motor actions. Admittedly, however, the neuronal sites and mechanisms underlying free will in the high cerebral centers are still an enigma (
Wegner, 2002
).
One of the major ideas that Sherrington outlined in his 1906 book “The Integrative Action of the Nervous System” was that complex actions of the nervous system could be composed of a collation of reflexes, somewhat like building a house by piling up bricks. From the control systems perspective, there now appear to be at least eight ways to integrate reflexes into the overall control of movement. First, many that are driven by different sensory inputs may share the same controller and controlled object (
Figure 9A
). For example, three types of relatively slow ocular reflexes are driven individually by vestibular or visual stimuli, as will be seen later in
Chapter 10
, “
Ocular Reflexes
.” Nonetheless, they commonly share vestibular nuclear neurons as the controller, and eyeballs and the associated oculomotor system, as the controlled object. By this means, such a group of reflexes can achieve the common purpose of securing visual stability and acuity under natural behavioral conditions. In other words, these individual ocular reflexes are combined together to form a “multi-input” control system. Second, several individual reflexes may have different controllers (
Figure 9B
, Reflex 1, 2, 3 controllers), but they may share the same controlled object. For example, a slow ocular reflex can be integrated with a brisk saccade only in the form of half-fused control because these eye movements require controllers having substantially different properties for generating slow and brisk eye movements, respectively (
Chapter 10
). Third, reflexes may also be combined with a voluntary motor control system in a hybrid way (
Figure 9C
)
because of the similarity of control system structures for reflexes and voluntary movements (see
Section 6
). Design problems in such hybrid systems will be discussed later (
Chapter 15
).
Figure 9. Schematics of types of integrated reflex control.
(A) A multi-input reflex. (B) Half-fused system. (C) Hybrid control of a reflex and a voluntary movement. For further explanation, see the text. Abbreviations: I
1,2,3
, three sensory inputs of different modality; VM, voluntary movement.

The spinal cord and brainstem contain a collation of reflexes to elaborate compound movements such as when assuming a posture, or when walking, swimming, and flying. Hence, a fourth way of integration is for some reflexes to be combined by mutual interaction (
Figure 10A
, Reflexes 1 and 2). For example, when one explores the visible world, a saccade and a head movement, the latter inducing the vestibuoocular reflex, occur in combination. This eye-head coordination involves an inhibitory cross talk between the independent eye and head controllers (
Kardamakis and Moschovakis, 2009
). A fifth way is for reflexes to be compounded when signals in a descending tract activate some combinations of reflexes to express behaviorally meaningful compound reflexes (
Figure 10B
, for review, see
Lemon, 2008
). Anders Lundberg (1920–2009) and his colleagues found a good example in the cervical segments of the spinal cord. In the C3–C4 propriospinal system of the cat, interneurons were shown to receive extensive convergence from different primary sensory afferents and supraspinal centers (
Lundberg, 1999
;
Alsternark et al., 2007
). Through excitation or inhibition of relevant interneurons in this system, signals of each descending tract could produce compound reflexes to provide desired movement patterns, such as target reaching by the hand (
Chapter 13
). Operation of segmental spinal circuits sometimes involves a type of function generator (FG in
Figure 10B
,
b
). Locomotion is a good example of this type of compounding reflex. It involves flexion reflexes, crossed extension reflexes, interlimb coordination, and, in addition, a central pattern generator (CPG) mechanism for rhythm generation (
Grillner et al., 1991
,
2007
;
Grillner and Jessel, 2009
) (see
Chapter 11
).
Figure 10. Schematics of the mutual interaction, compounding, and neuromodulation of reflexes.
Arrows denote synaptic actions, either excitatory or inhibitory. A shows how reflexes are combined by mutual interactions (Reflexes 1 and 2). In B, the compounding of reflexes 1, 3, and 4 is brought about by commands from descending tracts (a). Alternatively, reflexes 2, 3, and 4 can be coactivated by descending tracts (b) via FG, a function-generator. C shows how reflexes are modulated by aminergic and/or peptidergic innervation (represented by m and n) to exhibit a specific pattern of combination for behavior (m to reflexes 1, 2, and 4 and n to reflexes 1, 3, and 4). “Incentive” means a stimulus that leads to a specific behavior.
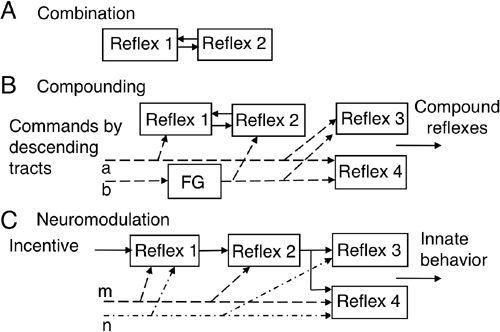
The sixth way to integrate reflexes is by “neuromodulation.” As demonstrated in the crustacean stomatogastric nervous system (
Marder et al., 1986
;
Selverston 1995
), a small amount of a single peptide or amine may instantaneously rewire a neuronal circuit and switch behavior expression of the system. This mechanism may apply to the hypothalamus located in the most rostral and ventral part of the brainstem, which regulates innate behaviors including food intake, drinking, and reproduction; they are evoked by incentives such as the need for food, water, and reproductive activity, respectively. These behaviors involve a series of complex movements in order to approach and acquire the incentives. On the other hand, noxious stimuli such as drinking stale water or the figure of an enemy induce aversion, aggression, or defense reactions. An innate behavior involves a combination of reflexes and compound movements. For example, food intake involves locomotion to approach the food, rhythmic mastication, and the swallowing reflex. The hypothalamus contains a number of innate behavioral centers, each of which produces a specific pattern of behavior by secreting a neuromodulator substance through their widely distributed axons in the brainstem and spinal cord. The secreted neuromodulator substance may activate or inhibit a number of component reflexes and compounded movements (
Figure 10C
); hence, one circuit can be configured to perform a variety of different behaviors by activating neurons via certain types of neuromodulator receptors.
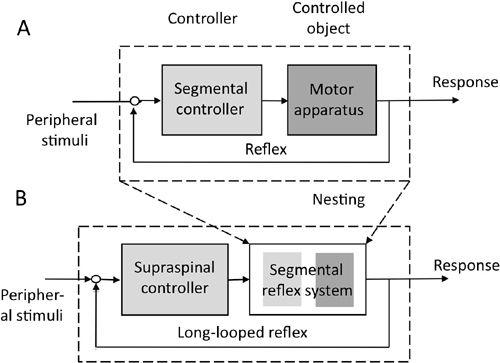
The seventh way to use reflexes, and probably the most important in regard to the cerebellum, is by “nesting” (as in “matryoshka”), which has been used to explain perceptual organization (
Leyton, 1987
) and even the entire hierarchical organization of the CNS (
Baev, 1999
). The nesting idea is that a reflex composed of a controller and controlled object at the lowest level can be regarded as a controlled object at the next higher level. For example, a stretch reflex is a control system at the segmental level (
Figure 11A
), but at a brainstem level, it acts as the controlled object of the vestibulospinal descending tract neurons, which act as the controller (
Figure 11B
). In a similar vein, the primary motor cortex acts as a controller of the spinal segmental circuits, which are the controlled objects (
Figure 12A
,
B
). Furthermore, the entire corticospinal system constitutes a controlled object for the premotor cortex, which serves as its controller (
Figure 12C
). Through use of this nesting principle, collective reflexes integrated in the previous six ways constitute a controlled object for a higher-level controller, which can thereby exert control over many reflexes in various combinations.
Figure 11. The one-step nesting of a reflex within a supraspinal controller.
(A) The control system structure of a segmental reflex. (B) A being nested within the controlled object of a supraspinal controller. For further details, see the text.