Resurrecting Pompeii (39 page)
Read Resurrecting Pompeii Online
Authors: Estelle Lazer

Additional cranial material that could be used for comparison with the Pompeian sample was less satisfactory and included an assortment of European populations, which dated from the prehistoric to the modern period. In addition, where data were available, a sample of Ancient Egyptian skulls, prehistoric Africans from Mali and an historic Nubian sample were compared with the Pompeian material as an acknowledgement of the possibility of Pompeian contact with Africa.
18
As there was very little published data for the population incidence of double-rooted canines and post-cranial nonmetric traits, comparisons were made with all the available material.
A major consideration in relation to comparison of traits with other published material is the lack of standardization in the presentation of trait incidence. Scholars have not always described the techniques that they used to record incidence. There are further problems when making comparisons between data sets for different populations, which include differences in sample sizes and whether they reflect random samples or are representative of the populations from which they were drawn. Apart from the acknowledgement that they exist, it is very difficult to account for these problems.
19
The samples of victims of the
AD
79 eruption are particularly problematic as comparative material since they reflect a sample of disaster victims, which is unusual in the archaeological record. The fact that they share roughly the same date and cause of death means that they provide more of a snapshot than a very slow time lapse view of a population. Statistical analyses were used to identify whether there was any association between the traits, with sex, or sides for bilateral features.
20
The results for the majority of these traits were inconclusive. This is partly attributable to the lack of appropriate comparative material, though investing meaning in the patterns of expression of non-metric traits is problematic (see below). The non-metric traits that yielded the most interesting results in relation to the issue of heterogeneity in the Pompeian cranial sample were palatine torus and double-rooted canines. While perhaps not as significant as population indicators for the Pompeian sample, it is also worth considering some of the other Pompeian cranial traits that can be compared with the data obtained by Nicolucci and Capasso. These include the expression of the metopic suture, wormian or fontanelle bones and inca bones and frontal grooves.
This is a hyperostotic trait, which means that it is associated with excessive ossification into structures that are usually made up of cartilage or dura.
21
Palatine torii can be observed on the palate as a median, or more frequently, paramedian bony mound, varying in height, width and length. They tend to be bilateral but can be unilateral. There can be a considerable range of expression of this trait from very slight and more pronounced at either the anterior or posterior end of the bone to excessive and covering the entire length of the palate. Researchers disagree about the relationship of age and expression of palatine torus and there are huge discrepancies between reports of first appearance and when development ceases. Similarly, there is no consensus about its aetiology. Some scholars have suggested that this is a threshold trait, which means that expression is dependent on environmental factors reaching a particular level. Stress from mastication has been invoked as a contributing factor to the development of palatine torus. Higher frequencies have been observed in circumpolar populations, whose diets include frozen and dried meat, which require hard chewing. It has also been suggested that nutrition may have some impact on the appearance of this trait. The consumption of marine food, with elevated levels of omega-3 and omega-6 fatty acids as well as vitamin D, has been proposed as major contributor to the development of palatine torus. Circumpolar populations have again been cited to support this notion as they tend to rely on a marine diet. Familial studies indicate that there appears to be a genetic potential for the expression of palatine torus. Observed occurrence of this trait in monozygotic twins further supports a genetic element, though it has been suggested that its presence can be influenced by other factors, such as pathology. It is most likely that palatine torus results from the interaction between genetic and environmental components.
22
Some degree of expression for palatine torus was present on all but two cases of the 52 skulls in the Forum Bath collection where the palate had been preserved. A similar frequency was observed in the Sarno Bath collection, though the survival rate of the palate was much lower in this collection. Contrary to the literature on palatine torus,
23
this trait was not found to be associated with either sex or age in the Pompeian sample. The Pompeian incidence for palatine torus is 96.2 per cent, which is extremely high compared to that recorded for most other populations (Table 9.1). Only three cases of palatine torus were reported for the Herculaneum sample studied by Capasso.
24
He did not mention the sample size for his observations of this trait, but based on the frequency calculations of the other epigenetic features he documented, this apparently represents 1.9 per cent of his sample.
It is notable that high frequencies have been reported for this trait in populations that are very genetically distant from the Pompeian sample. For example, Pardoe reported an incidence of palatine torus of up to 74.5 per cent for Australian Aboriginal samples from the border area between New South Wales and Victoria.
25
Even higher frequencies have been recorded for archaeological samples of circumpolar populations (Table 9.2).
The frequency of palatine torus in the Pompeian sample is extraordinary in comparison with the majority of the available data for Italian and other Mediterranean populations, especially that from Herculaneum, and requires some investigation. It is important to note that the case of 100 per cent expression from Termoli is based on a sample size of only nine individuals. It could be argued that the high frequency is an artefact of the comparatively low survival rate of this portion of the skull. There is, however, no reason to assume that preservation is not random. Even if it were strongly skewed toward individuals with this trait, the frequency would still be higher than that for most of the comparative material. In addition, observations of palatine torus for the available Sarno Bath palate sample, which were not included in this study, were consistent with the documented frequency for the Forum Bath collection.
Consideration should also be given to scoring. Berry and Berry claimed that there was a discrepancy between observers in scoring this trait as the results obtained varied considerably from other published data. This led them to postulate the existence of two separate entities that could be scored as palatine torus.
26
This problem should have been solved by the publication of Hauser and De Stefano’s standard atlas in 1989, which includes photographic references to minimize ambiguity for each trait. Another source of possible over-identification of this trait in comparison to the results of other scholars is the inclusion of trace scores. Some observers score trace expression
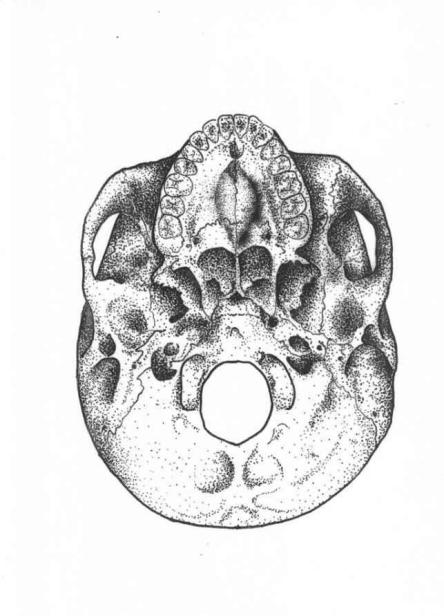
Figure 9.3
Basilar view of skull, showing a strongly expressed palatine torus on the bony palate (cf. Fig. A3.4 where this trait is absent) (adapted from Comas, 1960, in Krogman, 1962, 318, and Brothwell, 1981, 94; Hauser and Stefano, 1989, 175–7)
Table 9.1
Presence of palatine torus in various populations
Pompeii
AD
79 52 96.2
Pompeii
AD
79 (trace cases excluded) 52 73.1
Herculaneum
AD
79 159 1.9
Pontecagnano (Campania) (7
th
–6
th
century
BC
) 32 21.9
Sala Consilina (Campania) (9
th
–6
th
century
BC
)9 0 Termoli (Molise) (7
th
century
BC
) 9 100 Ardea (Latium) (8
th
– 6
th
century
BC
)190 Romans (Latium) (6
th
–5
th
century
BC
) 167 14.9
Alfedena (Abruzzo) (6
th
century
BC
) 69 34.8
Campovalano (Abruzzo) (7
th
–6
th
century
BC
) 95 32.7
Perdasdefogu (Sardinia) (9
th
century
BC
)17 0 Etruscans 1 (Central Etruria) (6
th
–5
th
century
BC
)56 0 Etruscans 2 (Southern Etruria) (6
th
–5
th
century
BC
)84 0 San Vincenzo al Volturno 153 23.7
Cefalu (17
th
century
BC
) 13 7.7
Plemmyrion (16
th
–14
th
century
BC
)333 Castiglione (17
th
century
BC
) 11 18.2
Castiglione (8
th
– 6
th
century
BC
) 6 33.3
Thapsos (16
th
–14
th
century
BC
) 41 2.4
Lentini (5
th
– 4
th
century
BC
) 6 33.3
Siracusa (8
th
century
BC
) 29 6.9
Siracusa (3
rd
century
BC
) 109 11.9
Piscitello (5
th
– 4
th
century
BC
)270 Carlentini (5
th
– 4
th
century
BC
)110 Modern Roman sample 285 7.4
Undated Sardinian population 245 7.3
African sample (Mali) (1
st
millennium
BC
) 145 0 Nubian (historic) 33 3
Sources: Adapted from Capasso, 2001, 982; Hauser and De Stefano, 1989, 178 –79; V. Higgins (University of Notre Dame, Rome) to E. Lazer, 1989–1990, personal communication; Lazer, 1995, 297; Rubini
et al
., 1999, 10; Rubini
et al
., 2007, 124.
Frequency of palatine torus from Scandinavian archaeological samples
Population Sample size Frequency (%)
Medieval Norway (male)
Medieval Norway (female)
Medieval Iceland (male)
Medieval Iceland (female)
Eastern Early Greenland (male)
Eastern Early Greenland (female) Eastern Middle–Late Greenland (male) Eastern Middle–Late Greenland (female) Western Greenland (male)
Western Greenland (female)
48 70.8 50 90.0 20 75.0 34 91.2 20 80.0 11 90.9 11 90.9 17 100
et al
., 1992, 151.
as absent.
27
Trace expression only accounts for 12 cases or 23.1 per cent of the Pompeian sample. Removal of all cases with a trace score still leaves an unequivocal 73.1 per cent with palatine torus. Since Capasso used the same standard scoring system as the Pompeii study, the lower frequency reported for the Herculanuem sample cannot be attributed to differences in recording. Nonetheless, it would be valuable for other ancient and contemporary skulls from the Vesuvian region to be examined for palatine torus to establish if this is a feature that is specific to ancient Pompeii or whether there are other populations in the region where the incidence of expression is high.
It does appear that the high frequency of this trait in the Pompeian sample is not an artefact. As already mentioned, the aetiology of this trait is not well understood. It appears that both environmental and genetic components contribute to the expression of palatine torus. Whatever the mechanism for the formation of the palatine torus, it has the potential to be a useful population descriptor for ancient Pompeians. It could be argued that the almost total presence of the trait in the sample might suggest a type of homogeneity that was not necessarily based on similarity of genotype but perhaps a shared environment during the period of osseous development. Dietary factors should be considered, though it would be hard to explain why there is such a low prevalence of this trait in the nearby settlement of Herculaneum, which has been argued to have relied heavily on marine protein (Chapter 8).
The roots of canines in the mandible or lower jaw are occasionally divided into two parts: labial (facing the lips) and lingual (facing toward the tongue). The degree of division can vary and be either partial or complete. It is most uncommon to find a bifurcated root on an upper or maxillary canine.
28
This characteristic can be a useful population marker. During the course of research, loose canines were routinely removed from their sockets to facilitate measurement and a number of double-rooted canines were observed. Observations were also made of the sockets of canines that had been lost postmortem. This is only useful for the identification of this trait when the roots are well divided. Scoring was limited to unequivocal cases. Only mandibular occurrence of this trait was observed by these means. Six of the 21 mandibles from which it was possible to make observations had teeth with roots that were divided. It is perhaps misleading to use percentages for such a small sample size, but for the purposes of comparison, the prevalence of doublerooted canines was about 28.6 per cent.
There is minimal comparative data for this trait in the literature. Maxillary canines of 13 Central Southern Italian Iron Age populations from either side of the Apennine Mountains and dating from the ninth to the second centuries
BC
were examined for the presence of bifurcated roots. Not surprisingly, no cases were observed in the 1,114 individuals, which included three Campanian samples from the region to the south of Naples (Coppa
et al
. 1998: 375). The mandibular canines were not examined for this trait. Scott and Turner observed double-rooted canines in varying frequencies in a diverse sample of populations. Table 9.3 demonstrates that this trait is extremely rare in Asiatic, Oceanic and African populations. Turner found double-rooted canines to occur more frequently in European populations.
29
Because of the small sample size and the lack of appropriate comparative data, it is not reasonable to draw too many conclusions from the presence of double-rooted canines in the Pompeian sample. The frequency appears to be considerably higher for Pompeians than for any other recorded population. This finding does appear to be remarkable and should be investigated further. The entire Pompeian skeletal sample should be subject to more detailed examination, with x-ray analysis of the jaws which still had teeth in-situ that could not be removed for inspection, as well as the cast collection. The Herculaneum skeletal collection should also be inspected for this trait, which, as yet, has not been recorded in the sample, as well as other ancient and modern samples from Campania and the rest of Italy to establish whether this feature is unique to the site of Pompeii or whether it has a high regional occurrence.