In Search of Memory: The Emergence of a New Science of Mind (40 page)
Read In Search of Memory: The Emergence of a New Science of Mind Online
Authors: Eric R. Kandel
Tags: #Psychology, #Cognitive Psychology & Cognition, #Cognitive Psychology

The biotechnology industry now stands as a parallel career pathway for both young and mature scientists. Since the quality of science at the best companies is very high, it is likely that scientists will move freely between academic science and the biotechnology industry.
While the emergence of Memory Pharmaceuticals and other biotechnology companies has bolstered the hope of alleviating memory loss and created new career paths for scientists working on the brain, it also has raised the ethical issue of cognitive enhancement. Is it desirable to improve memory in normal people? Would it be desirable for young people who could afford them to buy memory-enhancing drugs before taking the college entrance exams? There is a range of opinions on this issue, but mine is that
healthy
young people are capable of studying and learning on their own and in school without the aid of chemical memory enhancers (students with learning disabilities might be considered differently). Studying well is, without a doubt, the best cognitive enhancer for those capable of learning.
In a larger sense, these issues raise ethical questions parallel to those raised regarding gene cloning and stem cell biology. The biological community is working in areas in which honest and well-informed people disagree about the ethical implications of the products of research.
How do we link advances in science with an adequate discussion of the ethical implications of science? Here two issues converge. The first relates to scientific research. Freedom to do research is like free speech, and we as a democratic society should, within rather broad limits, protect the freedom of scientists to carry out research wherever it takes them. If we in the United States prohibit research in a particular area of science, we can be sure it will be done elsewhere, perhaps even in a part of the world where human life is not valued as highly or thought about as extensively as it is here. The second issue relates to an evaluation of how, if at all, a scientific discovery is to be used. This evaluation should not be left to scientists, because it affects society at large. Scientists can contribute to discussions about how the products of science are to be used, but final decisions require the participation of ethicists, lawyers, patients’ rights groups, and clergy, as well as scientists.
Ethics, a subfield of philosophy, historically has been concerned with the moral issues of mankind. Biotechnology gave rise to the specialized field of bioethics, which is concerned with the social and moral implications of biological and medical research. To address the particular issues raised by the new science of mind, William Safire, a columnist for
The New York Times
and president of the DANA Foundation, a public-interest group devoted to familiarizing the general public with the importance of brain science, encouraged the foundation in 2002 to stimulate studies in the field of neuroethics. To start this off, Safire sponsored a symposium entitled
Neuroethics: Mapping the Field
. The symposium brought together scientists, philosophers, lawyers, and clergy to address how the new view of mind affects issues that range from personal responsibility and free will to the competence of a mentally ill person to stand trial and the implications for society and the individual of new pharmacological modes of treatment.
To address the issues surrounding cognitive enhancers, I joined in 2004 with Martha Farah from the University of Pennsylvania, Judy llles from Stanford’s Center for Biomedical Ethics, Robin Cook-Deegan of the Center for Genome Ethics, Law and Policy of Duke University, and several other scholars. We published our declaration in
Nature Reviews Neuroscience
as a review article entitled “Neurocognitive Enhancement: What Can We Do and What Should We Do?”
The DANA Foundation is keeping an open discussion of neuroethical issues going. As Steven Hyman, the provost of Harvard University, put it in a recent DANA publication, “Matters…ranging from brain privacy to enhancement of mood and memory should be matters of vigorous discussion, and ideally those discussions will mature before continued scientific advances force societies to respond.”
J
ust as my studies of explicit memory in the 1990s had drawn me back to the issues that had attracted me to psychoanalysis in college, so the ability to study age-related memory disorders in mice at the beginning of the new millenium drew me irresistibly to the issues that had fascinated me as a resident in psychiatry. This renewed fascination with mental disorders was the result of several factors.
First, the biological research on memory that I was doing had progressed to the point that I could begin to address problems related to complex forms of memory and to the role of selective attention in memory, and this encouraged me to try to develop other animal models of mental illness. I was further attracted by the discovery that some forms of mental illnesses, such as post-traumatic stress disorders, schizophrenia, and depression, are accompanied by one or another type of memory impairment. As my understanding of the molecular biology of memory deepened and as I learned how instructive mouse models of age-related memory loss had proven to be, it became possible to think about the role of memory dysfunction in other forms of mental illness, and even in the biology of mental wellness.
Second, psychiatry had undergone a major shift toward biology in the course of my career. In the 1960s, when I was a resident at the Massachusetts Mental Health Center, most psychiatrists thought that the social determinants of behavior were completely independent of the biological determinants and that each acted on different aspects of mind. Psychiatric illnesses were classified into two major groups—organic illnesses and functional illnesses—based on presumed differences in their origin. That classification, which dated to the nineteenth century, emerged from postmortem examinations of the brains of mental patients.
The methods available for examining the brain at that time were too limited to detect subtle anatomical changes. As a result, only mental disorders that entailed significant loss of nerve cells and brain tissue, such as Alzheimer’s disease, Huntington’s disease, and chronic alcoholism, were classified as organic, or based in biology. Schizophrenia, the various forms of depression, and the anxiety states produced no loss of nerve cells or other obvious changes in brain anatomy and therefore were classified as functional, or not based in biology. Often, a special social stigma was attached to the so-called functional mental illnesses because they were said to be “all in a patient’s mind.” This notion was accompanied by the suggestion that the illness may have been put into the patient’s mind by his or her parents.
We no longer think that only certain diseases affect mental states through biological changes in the brain. Indeed, the underlying precept of the new science of mind is that
all
mental processes are biological—they all depend on organic molecules and cellular processes that occur literally “in our heads.” Therefore, any disorder or alteration of those processes must also have a biological basis.
Finally, I was asked in 2001 to write a paper for the
Journal of the American Medical Association
about molecular biological contributions to neurology and psychiatry with Max Cowan, a longtime friend who was vice president and senior scientific officer of the Howard Hughes Medical Institute. In writing the review, I was struck by the radical way in which molecular genetics and animal models of disease had transformed neurology, but had not psychiatry. This led me to wonder why molecular biology has not had a similar transformative effect on psychiatry.
The fundamental reason is that neurological diseases and psychiatric diseases differ in several important ways. Neurology has long been based on the knowledge of where in the brain specific diseases are located. The diseases that form the central concern of neurology—strokes, brain tumors, and the degenerative diseases of the brain—produce clearly discernible structural damage. Studies of those disorders taught us that, in neurology, location is key. We have known for almost a century that Huntington’s disease is a disorder of the caudate nucleus of the brain, Parkinson’s disease is a disorder of the substantia nigra, and amyotrophic lateral sclerosis (ALS) is a disorder of motor neurons. We know that each of these diseases produces its distinctive disturbances of movement because each involves a different component of the motor system.
In addition, a number of common neurological illnesses, such as Huntington’s, the fragile X form of mental retardation, some forms of ALS, and the early onset form of Alzheimer’s, were found to be inherited in a relatively straightforward way, implying that each of these diseases is caused by a single defective gene. Pinpointing the genes that produce these diseases has been relatively easy. Once a mutation is identified, it becomes possible to express the mutant gene in mice and flies and thus to discover how the gene gives rise to disease.
As a result of knowing the anatomical location, the identity, and the mechanism of action of specific genes, physicians no longer diagnose neurological disorders solely on the basis of behavioral symptoms. Since the 1990s, in addition to examining patients in the office, physicians can order tests for the dysfunction of specific genes, proteins, and nerve cell components, and they can examine brain scans to see how specific regions have been affected by a disorder.
TRACING THE CAUSES OF MENTAL ILLNESS IS A MUCH MORE
difficult task than locating structural damage in the brain. A century of postmortem studies of the brains of mentally ill persons failed to reveal the clear, localized lesions seen in neurological illness. Moreover, psychiatric illnesses are disturbances of higher mental function. The anxiety states and the various forms of depression are disorders of emotion, whereas schizophrenia is a disorder of thought. Emotion and thinking are complex mental processes mediated by complex neural circuitry. Until quite recently, little was known about the neural circuits involved in normal thought and emotion.
Furthermore, although most mental illnesses have an important genetic component, they do not have straightforward inheritance patterns, because they are not caused by mutations of a single gene. Thus, there is no single gene for schizophrenia, just as there is no single gene for anxiety disorders, depression, or most other mental illnesses. Instead, the genetic components of these diseases arise from the interaction of several genes with the environment. Each gene exerts a relatively small effect, but together they create a genetic predisposition—a potential—for a disorder. Most psychiatric disorders are caused by a combination of these genetic predispositions and some additional, environmental factors. For example, identical twins have identical genes. If one twin has Huntington’s disease, so will the other. But if one twin has schizophrenia, the other has only a 50 percent chance of developing the disease. To trigger schizophrenia, some other, non-genetic factors in early life—such as intrauterine infection, malnutrition, stress, or the sperm of an elderly father—are required. Because of this complexity in the pattern of inheritance, we have not yet identified most of the genes involved in the major mental illnesses.
In moving from implicit memory in
Aplysia
to explicit memory and the internal representation of space in the mouse, I had moved from a relatively simple realm to a far more complex one, a realm that held many questions of broad significance for human behavior but few solid insights. In trying to explore animal models of mental disorders I was taking a further step into uncertainty. Moreover, whereas I had been early in the study of implicit memory in
Aplysia
and had entered at an interesting midpoint in the study of explicit memory in the mouse, I was a late entry into the biology of mental disorders. Many other people had worked on animal models of mental disorders before me.
Lack of knowledge about the anatomy, genetics, and neural circuitry involved in mental disorders made it difficult to model them in animals. The one clear exception, and the one I focused on initially, was anxiety states. It is difficult to know whether a mouse ever suffers from schizophrenia, whether it is deluded or hallucinating. It is similarly difficult to recognize a mouse that is psychotically depressed. But every animal with a well-developed central nervous system—from snails to mice to monkeys to people—can become afraid, or anxious. In addition, fear has distinctive, easily recognizable features in each of these animals. Thus, not only do animals experience fear, but we can tell when they are anxious. We can, so to speak, read their thoughts. This insight was first set out by Charles Darwin in his classic 1872 study
The Expression of the Emotions in Man and Animals
.
The key biological fact that Darwin appreciated, and that has facilitated the development of animal models of anxiety states, is that anxiety—fear itself—is a universal, instinctive response to a threat to one’s body or social status and is therefore critical for survival. Anxiety signals a potential threat, which requires an adaptive response. As Freud pointed out, normal anxiety contributes to the mastery of difficult situations and thus to personal growth. Normal anxiety exists in two major forms: instinctive anxiety (instinctive or innate fear), which is built into the organism and is under more rigid genetic control, and learned anxiety (learned fear), to which an organism may be genetically predisposed but which is basically acquired through experience. As we have seen, instinctive anxiety can easily become associated through learning with a neutral stimulus. Since any capability that enhances survival tends to be conserved through evolution, both instinctive and learned fear are conserved throughout the animal kingdom (figure 25–1).
Both forms of fear can be deranged. Instinctive anxiety is pathological when it is excessive and persistent enough to paralyze action. Learned anxiety is pathological when it is provoked by events that present no real threat, as when a neutral stimulus comes to be associated in the brain with instinctive anxiety. Anxiety states were of particular interest to me because they are by far the most common mental illnesses: at some point in their lives, 10 to 30 percent of people in the general population suffer from these anxiety disorders!
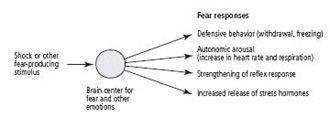
25–1
Defensive responses to fear that have been conserved through evolution.
By studying instinctive and learned fear in people and in experimental animals, we have gained much insight into both the behavioral and the biological mechanisms of instinctive and learned fear in people. One of the first behavioral insights was stimulated by the theories of Freud and the American philosopher William James, who realized that fear has both conscious and unconscious components. What was not clear was how the two components interact.
Traditionally, fear in people was thought to begin with conscious perception of an important event, such as seeing one’s house on fire. This recognition produces in the cerebral cortex an emotional experience—fear—that triggers signals to the heart, blood vessels, adrenal glands, and sweat glands to mobilize the body in preparation for defense or escape. Thus, according to this view, a conscious, emotional event initiates the later unconscious, reflexive, and autonomic defensive responses in the body.
James rejected this view. In a highly influential article published in 1884 and entitled “What Is Emotion?” he proposed that the cognitive experience of emotion is secondary to the physiological expression of emotion. He suggested that when we encounter a potentially dangerous situation—for example, a bear sitting in the middle of our path—our evaluation of the bear’s ferocity does not generate a consciously experienced emotional state. We do not experience fear until after we have run away from the bear. We first act instinctively and then invoke cognition to explain the changes in the body associated with that action.
Based on this idea, James and the Danish psychologist Carl Lange proposed that the conscious experience of emotion occurs only
after
the cortex has received signals about changes in one’s physiological state. In other words, conscious feelings are preceded by certain unconscious physiological changes—an increase or decrease in blood pressure, heart rate, and muscular tension. Thus, when you see a fire, you feel afraid because your cortex has just received signals about your racing heart, knocking knees, and sweaty palms. James wrote: “We feel sorry because we cry, angry because we strike, afraid because we tremble, and not that we cry, strike or tremble because we are sorry, angry or fearful, as the case may be.” According to this view, emotions are cognitive responses to information from bodily states mediated in good part by the autonomic nervous system. Our everyday experience confirms that information from the body contributes to emotional experience.
Experimental evidence soon supported some aspects of the James-Lange theory. For example, objectively distinguishable emotions are correlated with specific patterns of autonomic, endocrine, and voluntary responses. Furthermore, people whose spinal cord has been accidentally severed, cutting off feedback from the autonomic nervous system in regions of the body below the injury, appear to experience less intense emotions.