Why Evolution Is True (15 page)
Read Why Evolution Is True Online
Authors: Jerry A. Coyne

One of my favorite cases of embryological evidence for evolution is the furry human fetus. We are famously known as “naked apes” because, unlike other primates, we don’t have a thick coat of hair. But in fact for one brief period we do—as embryos. Around sixth months after conception, we become completely covered with a fine, downy coat of hair called
lanugo.
Lanugo is usually shed about a month before birth, when it’s replaced by the more sparsely distributed hair with which we’re born. (Premature infants, however, are sometimes born with lanugo, which soon falls off.) Now, there’s no need for a human embryo to have a transitory coat of hair. After all, it’s a cozy 98.6 degrees Fahrenheit in the womb. Lanugo can be explained only as a remnant of our primate ancestry: fetal monkeys also develop a coat of hair at about the same stage of development. Their hair, however, doesn’t fall out, but hangs on to become the adult coat. And, like humans, fetal whales also have lanugo, a remnant of when their ancestors lived on land.
lanugo.
Lanugo is usually shed about a month before birth, when it’s replaced by the more sparsely distributed hair with which we’re born. (Premature infants, however, are sometimes born with lanugo, which soon falls off.) Now, there’s no need for a human embryo to have a transitory coat of hair. After all, it’s a cozy 98.6 degrees Fahrenheit in the womb. Lanugo can be explained only as a remnant of our primate ancestry: fetal monkeys also develop a coat of hair at about the same stage of development. Their hair, however, doesn’t fall out, but hangs on to become the adult coat. And, like humans, fetal whales also have lanugo, a remnant of when their ancestors lived on land.
The final example from humans takes us into the realm of speculation, but is too appealing to omit. This is the “grasping reflex” of newborn babies. If you have access to an infant, gently stroke the palms of its hands. The baby will show a reflex response by making a fist around your finger. In fact, the grasp is so tight that an infant can, using both hands, hang for several minutes from a broomstick. (Warning: don’t try this experiment at home!) The grasping reflex, which disappears several months after birth, may well be an atavistic behavior. Newborn monkeys and apes have the same reflex, but it persists throughout the juvenile stage, allowing the young to hang on to their mother’s fur as they’re carried about.
It is sad that while embryology provides such a gold mine of evidence for evolution, textbooks of embryology often fail to point this out. I have met obstetricians, for instance, who know everything about the lanugo except why it appears in the first place.
As well as peculiarities of embryonic development, there are also peculiarities of animal structure that can be explained only by evolution. These are cases of “bad design.”
Bad DesignIN THE OTHERWISE FORGETTABLE MOVIE
Man of the
Year, comedian Robin Williams plays a television talk-show host who, through a series of bizarre accidents, becomes president of the United States. During a preelection debate, Williams’s character is asked about intelligent design. He responds, “People say intelligent design—we must teach intelligent design. Look at the human body; is that intelligent? You have a waste processing plant next to a recreation area!”
Man of the
Year, comedian Robin Williams plays a television talk-show host who, through a series of bizarre accidents, becomes president of the United States. During a preelection debate, Williams’s character is asked about intelligent design. He responds, “People say intelligent design—we must teach intelligent design. Look at the human body; is that intelligent? You have a waste processing plant next to a recreation area!”
It’s a good point. Although organisms appear designed to fit their natural environments, the idea
of perfect
design is an illusion. Every species is imperfect in many ways. Kiwis have useless wings, whales have a vestigial pelvis, and our appendix is a nefarious organ.
of perfect
design is an illusion. Every species is imperfect in many ways. Kiwis have useless wings, whales have a vestigial pelvis, and our appendix is a nefarious organ.
What I mean by “bad design” is the notion that if organisms were built from scratch by a designer—one who used the biological building blocks of nerves, muscles, bone, and so on—they would not have such imperfections. Perfect design would truly be the sign of a skilled and intelligent designer. Imperfect design is the mark of evolution; in fact, it’s precisely what we expect from evolution. We’ve learned that evolution doesn’t start from scratch. New parts evolve from old ones, and have to work well with the parts that have already evolved. Because of this, we should expect compromises: some features that work pretty well, but not as well as they might, or some features-like the kiwi wing—that don’t work at all, but are evolutionary leftovers.
A good example of bad design is the flounder, whose popularity as an eating fish (Dover sole, for instance) comes partly from its flatness, which makes it easy to bone. There are actually about five hundred species of flatfish—halibut, turbot, flounders, and their kin—all placed in the order Pleuronectiformes. The word means “side-swimmers,” a description that’s the key to their poor design. Flatfish are born as normal-looking fish that swim vertically, with one eye placed on each side of a pancake-shaped body. But a month thereafter, a strange thing happens: one eye begins to move upward. It migrates over the skull and joins the other eye to form a pair of eyes on one side of the body, either right or left, depending on the species. The skull also changes its shape to promote this movement, and there are changes in the fins and color. In concert, the flatfish tips onto its newly eyeless side, so that both eyes are now on top. It becomes a flat camouflaged bottom-dweller that preys on other fish. When it has to swim, it does so on its side. Flatfish are the world’s most asymmetrical vertebrates; check out a specimen the next time you go to the fish market.
If you wanted to design a flatfish, you wouldn’t do it this way. You’d produce a fish like the skate, which is flat from birth and lies on its belly-not one that has to achieve flatness by lying on its side, moving its eyes and deforming its skull. Flatfish are poorly designed. But the poor design comes from their evolutionary heritage. We know from their family tree that flounders, like all flatfish, evolved from “normal” symmetrical fish. Evidently, they found it advantageous to tip onto their sides and lie on the seafloor, hiding themselves from both predators and prey. This, of course, created a problem : the bottom eye would be both useless and easily injured. To fix this, natural selection took the tortuous but available route of moving its eye about, as well as otherwise deforming its body.
One of nature’s worst designs is shown by the recurrent laryngeal nerve of mammals. Running from the brain to the larynx, this nerve helps us speak and swallow. The curious thing is that it is much longer than it needs to be. Rather than taking a direct route from the brain to the larynx, a distance of about a foot in humans, the nerve runs down into our chest, loops around the aorta and a ligament derived from an artery, and then travels back up (“recurs”) to connect to the larynx (figure 19). It winds up being three feet long. In giraffes the nerve takes a similar path, but one that runs all the way down that long neck and back up again: a distance fifteen feet longer than the direct route! When I first heard about this strange nerve, I had trouble believing it. Wanting to see for myself, I mustered up my courage to make a trip to the human anatomy lab and inspect my first corpse. An obliging professor showed me the nerve, tracing its course with a pencil down the torso and back up to the throat.
This circuitous path of the recurrent laryngeal nerve is not only poor design, but might even be maladaptive. That extra length makes it more prone to injury. It can, for example, be damaged by a blow to the chest, making it hard to talk or swallow. But the pathway makes sense when we understand how the recurrent laryngeal nerve evolved. Like the mammalian aorta itself, it descends from those branchial arches of our fishlike ancestors. In the early fishlike embryos of all vertebrates, the nerve runs from top to bottom alongside the blood vessel of the sixth branchial arch; it is a branch of the larger vagus nerve that travels along the back from the brain. And in adult fish, the nerve remains in that position, connecting the brain to the gills and helping them pump water.
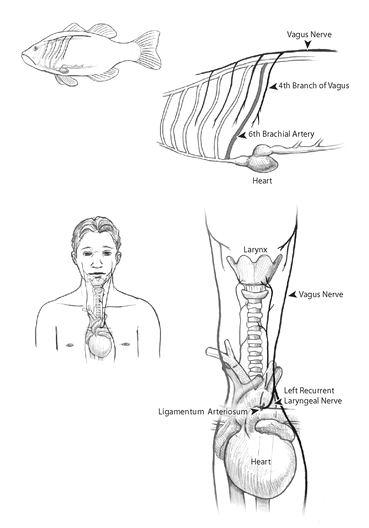
FIGURE 19
. The circuitous path of the left recurrent laryngeal nerve in humans is evidence for their evolution from a fishlike ancestor. In fish, the sixth branchial arch, which later becomes a gill, is served by the sixth aortic arch. The fourth branch of the vagus nerve runs behind this arch. These structures remain part of the gill apparatus in adult fish, innervating and bringing blood from the gills. In mammals, however, part of the branchial arch evolved into the larynx. The larynx and its nerve remained connected during this process, but the sixth aortic arch on the left side of the body moved down into the chest to become a nonfunctional remnant, the
ligamentum arteriosum.
Because the nerve remained behind this arch but still connected to a structure in the neck, it was forced to evolve a pathway that travels down into the chest, loops around the aorta and the remnants of the sixth aortic arch, and then travels back up to the larynx. The indirect path of this nerve does not reflect intelligent design but can be understood only as the product of our evolution from ancestors having very different bodies.
. The circuitous path of the left recurrent laryngeal nerve in humans is evidence for their evolution from a fishlike ancestor. In fish, the sixth branchial arch, which later becomes a gill, is served by the sixth aortic arch. The fourth branch of the vagus nerve runs behind this arch. These structures remain part of the gill apparatus in adult fish, innervating and bringing blood from the gills. In mammals, however, part of the branchial arch evolved into the larynx. The larynx and its nerve remained connected during this process, but the sixth aortic arch on the left side of the body moved down into the chest to become a nonfunctional remnant, the
ligamentum arteriosum.
Because the nerve remained behind this arch but still connected to a structure in the neck, it was forced to evolve a pathway that travels down into the chest, loops around the aorta and the remnants of the sixth aortic arch, and then travels back up to the larynx. The indirect path of this nerve does not reflect intelligent design but can be understood only as the product of our evolution from ancestors having very different bodies.
During our evolution, the blood vessel from the fifth arch disappeared, and the vessels from the fourth and sixth arches moved downward into the future torso so that they could become the aorta and a ligament connecting the aorta to the pulmonary artery. But the laryngeal nerve, still behind the sixth arch, had to remain connected to the embryonic structures that become the larynx, structures that remained near the brain. As the future aorta evolved backward toward the heart, the laryngeal nerve was forced to evolve backward along with it. It would have been more efficient for the nerve to detour around the aorta, breaking and then re-forming itself on a more direct course, but natural selection couldn’t manage that, for severing and rejoining a nerve is a step that reduces fitness. To keep up with the backward evolution of the aorta, the laryngeal nerve had to become long and recurrent. And that evolutionary path is recapitulated during development, since as embryos we begin with the ancestral fishlike pattern of nerves and blood vessels. In the end, we’re left with bad design.
Courtesy of evolution, human reproduction is also full of jerry-rigged features. We’ve already learned that the descent of male testes, a result of their evolution from fish gonads, creates weak spots in the abdominal cavity that can cause hernias. Males are further disadvantaged because of the poor design of the urethra, which happens to run right through the middle of the prostate gland that produces some of our seminal fluid. To paraphrase Robin Williams, it’s a sewage pipe running directly through a recreation area. A large fraction of males develop enlarged prostates later in life, which squeeze the urethra and make urination difficult and painful. (Presumably this wasn’t a problem during most of human evolution, when few men lived past thirty.) A smart designer wouldn’t put a collapsible tube through an organ prone to infection and swelling. It happened this way because the mammalian prostate gland evolved from tissue in the walls of the urethra.
Women don’t fare much better. They give birth through the pelvis, a painful and inefficient process that, before modern medicine, killed appreciable numbers of mothers and babies. The problem is that as we evolved a big brain, the infant’s head became very large relative to the opening of the pelvis, which had to remain narrow to allow efficient bipedal (two-legged) walking. This compromise leads to the difficulties and enormous pain of human birth. If you designed a human female, wouldn’t you have rerouted the female reproductive tract so it exited through the lower abdomen instead of the pelvis? Imagine how much easier it would be to give birth! But humans evolved from creatures that laid eggs or gave live birth—less painfully than we—through the pelvis. We’re constrained by our evolutionary history.
And would an intelligent designer have created the small gap between the human ovary and Fallopian tube, so that an egg must cross this gap before it can travel through the tube and implant in the uterus? Occasionally a fertilized egg doesn’t make the leap successfully and implants in the abdomen. This produces an “abdominal pregnancy,” almost invariably fatal to the baby and, without surgery, to the mother. The gap is a remnant of our fish and reptilian ancestors, who shed eggs directly from the ovary to the outside of their bodies. The Fallopian tube is an imperfect connection because it evolved later as an add-on in mammals.
20
20
Some creationists respond that poor design is not an argument for evolution—that a supernatural intelligent designer could nevertheless have created imperfect features. In his book
Darwin’s Black Box,
the ID proponent Michael Behe claims that “features that strike us as odd in a design might have been placed there by the Designer for a reason—for artistic reasons, for variety, to show off, for some as-yet-undetectable practical purpose, or for some unguessable reason—or they might not.” But this misses the point. Yes, a designer may have motives that are unfathomable. But the particular bad designs that we see make sense
only if they evolved from features of earlier ancestors.
If a designer did have discernible motives when creating species, one of them must surely have been to fool biologists by making organisms look as though they evolved.
Darwin’s Black Box,
the ID proponent Michael Behe claims that “features that strike us as odd in a design might have been placed there by the Designer for a reason—for artistic reasons, for variety, to show off, for some as-yet-undetectable practical purpose, or for some unguessable reason—or they might not.” But this misses the point. Yes, a designer may have motives that are unfathomable. But the particular bad designs that we see make sense
only if they evolved from features of earlier ancestors.
If a designer did have discernible motives when creating species, one of them must surely have been to fool biologists by making organisms look as though they evolved.
Chapter 4
The Geography of Life
When on board H.M.S. “Beagle” as naturalist,
I was much struck with certain facts in the distribution of
the inhabitants of South America, and in the geological
relations of the present to the past inhabitants of that continent.
These facts seemed to me to throw some light on the origin of
species—that mystery of mysteries, as it has been called
by one of our greatest philosophers.
I was much struck with certain facts in the distribution of
the inhabitants of South America, and in the geological
relations of the present to the past inhabitants of that continent.
These facts seemed to me to throw some light on the origin of
species—that mystery of mysteries, as it has been called
by one of our greatest philosophers.
On the Origin of Species
Other books
The Granite Moth by Erica Wright
Winds of Wyoming (A Kate Neilson Novel) by Lyles, Rebecca Carey
Zombie Day Care by Halloran, Craig
Murder in the Secret Garden by Ellery Adams
Attila the Hun by John Man
Rock All Night (The Rock Star's Seduction #2) by Olivia Thorne
Bury Her Deep by Catriona McPherson
A Hidden Secret by Linda Castillo
Ten Thousand Islands by Randy Wayne White
The Cinderella List by Judy Baer