Why Evolution Is True (14 page)
Read Why Evolution Is True Online
Authors: Jerry A. Coyne

And again, it was Darwin who reconciled the disparate facts about embryology that filled the textbooks of his time, and showed that the puzzling features of development suddenly made perfect sense under the unifying idea of evolution:
Embryology rises greatly in interest, when we thus look at the embryo as a picture, more or less obscured, of the common parent-form of each great class of animals.
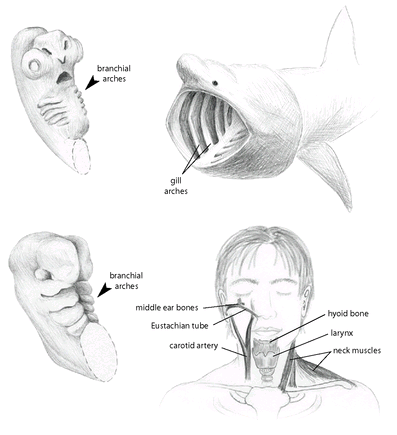
FIGURE 16
. Branchial arches of a shark embryo (top left) and a human embryo (bottom left). In sharks and fish (such as the basking shark
Cetorhinus maximus
shown at top right), the arches develop directly into the adult gill structures, while in the human (and other mammals) they develop into diverse structures in the adult head and upper body.
. Branchial arches of a shark embryo (top left) and a human embryo (bottom left). In sharks and fish (such as the basking shark
Cetorhinus maximus
shown at top right), the arches develop directly into the adult gill structures, while in the human (and other mammals) they develop into diverse structures in the adult head and upper body.
Let’s start with that fishy fetus of all vertebrates-limbless and sporting a fishlike tail. Perhaps the most striking fishlike feature is a series of five to seven pouches, separated by grooves, that lie on each side of the embryo near its future head. These pouches are called the
branchial
arches, but we’ll call them “arches” for short (figure 16). Each arch contains tissues that develop into nerves, blood vessels, muscles, and bone or cartilage. As fish and shark embryos develop, the first arch becomes the jaw and the rest become gill structures: the clefts between the pouches open up to become the gill slits, and the pouches develop nerves to control the movement of the gills, blood vessels to remove oxygen from water, and bars of bone or cartilage to support the gill structure. In fish and sharks, then, the development of gills from the embryonic arches is more or less direct: these embryonic features simply enlarge without much change to form the adult breathing apparatus.
branchial
arches, but we’ll call them “arches” for short (figure 16). Each arch contains tissues that develop into nerves, blood vessels, muscles, and bone or cartilage. As fish and shark embryos develop, the first arch becomes the jaw and the rest become gill structures: the clefts between the pouches open up to become the gill slits, and the pouches develop nerves to control the movement of the gills, blood vessels to remove oxygen from water, and bars of bone or cartilage to support the gill structure. In fish and sharks, then, the development of gills from the embryonic arches is more or less direct: these embryonic features simply enlarge without much change to form the adult breathing apparatus.
But in other vertebrates that don’t have gills as adults, these arches turn into very different structures-structures that make up the head. In mammals, for example, they form the three tiny bones of the middle ear, the Eustachian tube, the carotid artery, the tonsils, the larynx, and the cranial nerves. Sometimes the embryonic gill slits fail to close in human embryos, producing a baby with a cyst on its neck. This condition, an atavistic remnant of our fishy ancestors, can be corrected with surgery.
Our blood vessels go through especially strange contortions. In fish and sharks, the embryonic pattern of vessels develops without much change into the adult system. But as other vertebrates develop, the vessels move around, and some of them disappear. Mammals like ourselves are left with only three main vessels from the original six. The really curious thing is that as our development proceeds, the changes resemble an evolutionary sequence. Our fishlike circulatory system turns into one similar to that of embryonic amphibians. In amphibians, the embryonic vessels turn directly into adult vessels, but ours continue to change—into a circulatory system resembling that of embryonic reptiles. In reptiles, this system then develops directly into the adult one. But ours changes still further, adding a few more twists that turn it into a true mammalian circulatory system, complete with carotid, pulmonary, and dorsal arteries (figure 17).
These patterns raise a lot of questions. First, why do different vertebrates, which wind up looking very different from one another, all begin development looking like a fish embryo? Why do mammals form their heads and faces from the very same embryonic structures that become the gills of fish? Why do vertebrate embryos go through such a contorted sequence of changes in the circulatory system? Why don’t human embryos, or lizard embryos, begin development with their adult circulatory system already in place, rather than making a lot of changes in what developed earlier? And why does our sequence of development mimic the order of our ancestors (fish to amphibian to reptile to mammal)? As Darwin argued in
The Origin
, it’s not because human embryos experience a series of environments during development to which they must successively adapt—first a fishlike one, then a reptilian one, and so on:
The Origin
, it’s not because human embryos experience a series of environments during development to which they must successively adapt—first a fishlike one, then a reptilian one, and so on:
The points of structure, in which the embryos of widely different animals of the same class resemble each other, often have no direct relation to their conditions of existence. We cannot, for instance, suppose that in the embryos of the vertebrata the peculiar loop-like course of the arteries near the branchial slits are related to similar conditions—in the young mammal which is nourished in the womb of its mother, in the egg of the bird which is hatched in a nest, and in the spawn of a frog under water.
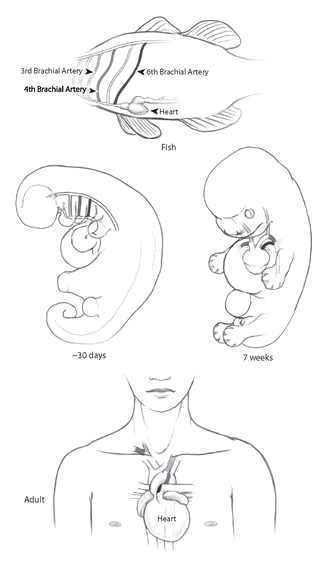
FIGURE 17
. The blood vessels of embryonic humans start out resembling those of embryonic fish, with a top and bottom vessel connected by parallel vessels, one on each side (“aortic arches”). In fish, these side vessels carry blood to and from the gills. Embryonic and adult fish have six pairs of arches; this is the basic ground plan that appears at the beginning of development of all vertebrates. In the human embryo, the first, second, and fifth arches form briefly at the beginning of development, but disappear by four weeks of age, when the third, fourth, and sixth arches (distinguished by different shades of gray) form. By seven weeks, the embryonic arches have rearranged themselves, looking much like the embryonic vessels of a reptile. In the final adult configuration, the vessels are rearranged still more, with some having vanished or transformed themselves into different vessels. The aortic arches of fish undergo no such transformation.
. The blood vessels of embryonic humans start out resembling those of embryonic fish, with a top and bottom vessel connected by parallel vessels, one on each side (“aortic arches”). In fish, these side vessels carry blood to and from the gills. Embryonic and adult fish have six pairs of arches; this is the basic ground plan that appears at the beginning of development of all vertebrates. In the human embryo, the first, second, and fifth arches form briefly at the beginning of development, but disappear by four weeks of age, when the third, fourth, and sixth arches (distinguished by different shades of gray) form. By seven weeks, the embryonic arches have rearranged themselves, looking much like the embryonic vessels of a reptile. In the final adult configuration, the vessels are rearranged still more, with some having vanished or transformed themselves into different vessels. The aortic arches of fish undergo no such transformation.
The “recapitulation” of an evolutionary sequence is seen in the developmental sequence of other organs—our kidneys, for example. During development, the human embryo actually forms three different types of kidneys, one after the other, with the first two discarded before our final kidney appears. And those transitory embryonic kidneys are similar to those we find in species that evolved before us in the fossil record—jawless fish and reptiles, respectively. What does this mean?
You could answer this question superficially as follows: each vertebrate undergoes development in a series of stages, and the sequence of those stages happens to follow the evolutionary sequence of its ancestors. So, for example, a lizard begins development resembling an embryonic fish, then somewhat later an embryonic amphibian, and finally an embryonic reptile. Mammals go through the same sequence, but add on the final stage of an embryonic mammal.
This answer is correct but only raises deeper issues. Why does development often occur in this way? Why doesn’t natural selection eliminate the “fish embryo” stage of human development, since a combination of a tail, fishlike gill arches, and a fishlike circulatory system doesn’t seem necessary for a human embryo? Why don’t we simply begin development as tiny humans—as some seventeenth-century biologists thought we did—and just get larger and larger until we’re born? Why all the transformation and rearrangement?
The probable answer—and it’s a good one—involves recognizing that as one species evolves into another, the descendant inherits the developmental program of its ancestor: that is, all the genes that form ancestral structures. And development is a very conservative process. Many structures that form later in development require biochemical “cues” from features that appear earlier. If, for example, you try to tinker with the circulatory system by remodeling it from the very onset of development, you might produce all sorts of adverse side effects in the formation of other structures, like bones, that mustn’t be changed. To avoid these deleterious side effects, it’s usually easier to simply tack some less drastic changes onto what is already a robust and basic developmental plan. It is best for things that
evolved
later to be programmed to
develop
later in the embryo.
evolved
later to be programmed to
develop
later in the embryo.
This “adding new stuff onto old” principle also explains why the sequence of developmental changes mirrors the evolutionary sequence of organisms. As one group evolves from another, it often adds its development program on top of the old one.
Noting this principle, Ernst Haecl
forms of ancestors. Human fetuses, for example, never resemble adult fish or reptiles, but in certain ways they do resemble embryonic fish and reptiles. Also, the recapitulation is neither strict nor inevitable: not every feature of an ancestor’s embryo appears in its descendants, nor do all stages of development unfold in a strict evolutionary order. Further, some species, like plants, have dispensed with nearly all traces of their ancestry during development. Haeckel’s law has fallen into disrepute not only because it wasn’t strictly true, but also because Haeckel was accused, largely unjustly, of fudging some drawings of early embryos to make them look more similar than they really are.
19
Yet we shouldn’t throw out the baby with the bathwater. Embryos still show a form of recapitulation: features that arose earlier in evolution often appear earlier in development. And this makes sense only if species have an evolutionary history.
forms of ancestors. Human fetuses, for example, never resemble adult fish or reptiles, but in certain ways they do resemble embryonic fish and reptiles. Also, the recapitulation is neither strict nor inevitable: not every feature of an ancestor’s embryo appears in its descendants, nor do all stages of development unfold in a strict evolutionary order. Further, some species, like plants, have dispensed with nearly all traces of their ancestry during development. Haeckel’s law has fallen into disrepute not only because it wasn’t strictly true, but also because Haeckel was accused, largely unjustly, of fudging some drawings of early embryos to make them look more similar than they really are.
19
Yet we shouldn’t throw out the baby with the bathwater. Embryos still show a form of recapitulation: features that arose earlier in evolution often appear earlier in development. And this makes sense only if species have an evolutionary history.
Now, we’re not absolutely sure why some species retain much of their evolutionary history during development. The “adding new stuff onto old” principle is just a hypothesis—an explanation for the facts of embryology. It’s hard to prove that it was easier for a developmental program to evolve one way rather than another. But the facts of embryology remain, and make sense only in light of evolution. All vertebrates begin development looking like embryonic fish because we all descended from a fishlil
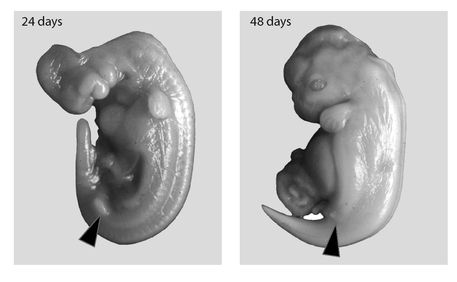
FIGURE 18
. The disappearing hindlimb structures in the spotted dolphin (
Stenella attenuata
)—evolutionary remnants of its four-legged ancestor. In the twenty-four-day-old embryo (left), the hindlimb bud (indicated by triangle) is well developed, only slightly smaller than the forelimb bud. By forty-eight days (right), the hindlimb buds have almost disappeared, while the forelimb buds continue to develop into what will be the flippers.
. The disappearing hindlimb structures in the spotted dolphin (
Stenella attenuata
)—evolutionary remnants of its four-legged ancestor. In the twenty-four-day-old embryo (left), the hindlimb bud (indicated by triangle) is well developed, only slightly smaller than the forelimb bud. By forty-eight days (right), the hindlimb buds have almost disappeared, while the forelimb buds continue to develop into what will be the flippers.
When he wrote
The Origin
, Darwin considered embryology his strongest evidence for evolution. Today he’d probably give pride of place to the fossil record. Nevertheless, science continues to accumulate intriguing features about development that support evolution. Embryonic whales and dolphins form hindlimb buds—bulges of tissue that, in four-legged mammals, become the rear legs. But in marine mammals the buds are reabsorbed soon after they’re formed. Figure 18 shows this regression in the development of the spotted dolphin. l3aleen whales, which lack teeth but whose ancestors were toothed whales, develop embryonic teeth that disappear before birth.
The Origin
, Darwin considered embryology his strongest evidence for evolution. Today he’d probably give pride of place to the fossil record. Nevertheless, science continues to accumulate intriguing features about development that support evolution. Embryonic whales and dolphins form hindlimb buds—bulges of tissue that, in four-legged mammals, become the rear legs. But in marine mammals the buds are reabsorbed soon after they’re formed. Figure 18 shows this regression in the development of the spotted dolphin. l3aleen whales, which lack teeth but whose ancestors were toothed whales, develop embryonic teeth that disappear before birth.
Other books
Sister of the Bride by Beverly Cleary
A Touch of Greed by Gary Ponzo
The Apprentices by Meloy, Maile
Cassidy Lane by Murnane, Maria
Dolores Claiborne by Stephen King
Packed and Ready to Go by Jacki Kelly
Rhythm of Us: Book 2 Of The Fated Hearts Series by Aimee Nicole Walker
Against a Brightening Sky by Jaime Lee Moyer
A Dog's Way Home by Bobbie Pyron
TW08 The Dracula Caper NEW by Simon Hawke