The Lying Stones of Marrakech (50 page)
Read The Lying Stones of Marrakech Online
Authors: Stephen Jay Gould

The uncovering of life's early history has featured several such cascades of discovery following a key insight about proper places to lookâand I introduce this year's wonderful story by citing a previous incident of remarkably similar character from the last generation of our science (literally so, for this year's discoverer wrote his Ph.D. dissertation under the guidance of the first innovator).
When, as a boy in the early 1950s, I first became fascinated with paleontology and evolution, the standard dogma about the origin of life proclaimed such an event inherently improbable, but achieved on this planet only because the immensity of geological time must convert the nearly impossible into the
virtually certain. (With no limit on the number of tries, you will eventually flip fifty heads in a row with an honest coin.) As evidence for asserting the exquisite specialness of life in the face of overwhelmingly contrary odds, these conventional sources cited the absence of any fossils representing the first half of the earth's existenceâa span of more than two billion years, often formally designated on older geological charts as the Azoic (literally “lifeless”) era. Although scientists do recognize the limitations of such “negative evidence” (the first example of a previously absent phenomenon may, after all, turn up tomorrow), this failure to find any fossils for geology's first two billion years did seem fairly persuasive. Paleontologists had been searching assiduously for more than a century and had found nothing but ambiguous scraps and blobs. Negative results based on such sustained effort over so many years do begin to inspire belief.
But the impasse broke in the 1950s, when Elso Barghoorn and Stanley Tyler reported fossils of unicellular life in rocks more than two billion years old. Paleontologists, to summarize a long and complex story with many exciting turns and notable heroes, had been looking in the wrong placeâin conventional sediments that rarely preserve the remains of single-celled bacterial organisms without hard parts. We had not realized that life had remained so simple for so long, or that the ordinary sites for good fossil records could not preserve such organisms.
Barghoorn and colleagues dispelled a century of frustration by looking in a different place, where cellular remains of bacteria might be preservedâin chert beds. Chert has the same chemical formula (with a different molecular arrangement) as quartz: silicon dioxide. Paleontologists rarely think of looking for fossils in silicate rocksâfor the perfectly valid and utterly obvious reason that silicates form by the cooling of volcanic magmas and therefore cannot contain organic remains. (Life, after all, doesn't flourish in bubbling lavas, and anything falling in gets burnt to a crisp.) But cherts can form at lower temperatures and be deposited amid layers of ordinary sediments in oceanic waters. Bacterial cells, when trapped in this equivalent of surrounding glass, can be preserved as fossils.
This cardinal insightâthat we had been searching in the wrong venue of ordinary (and barren) sediments rather than in fruitful chertsâcreated an entire field of study: collecting data for the first two-thirds or so of life's full history! Forty years later, we may look back with wonder at the flood of achievement and the complete overturn of established wisdom. We now possess a rich fossil record of early life, extending right back to the earliest potential source for cellular evidence. (The oldest rocks on earth that could preserve such data do
contain abundant fossils of bacterial organisms. These 3.5-to-3.6-billion-year-old rocks from Australia and South Africa are the most ancient strata on earth that have not been sufficiently altered by subsequent heat and pressure to destroy all anatomical evidence of life.)
Such ubiquity and abundance has forced a reversal of the old view. The origin of life at simplest bacterial grade now seems inevitable rather than improbable. As a mantra for memory, may I suggest: “Life on earth is as old as it could be.” I realize, of course, that an earliest possible appearance builds no proof for inevitability. After all, even a highly improbable event might occur, by good fortune, early in a series of trials. (You might flip those fifty successive heads on your tenth attemptâbut don't count, or bet, on it!) Nonetheless, faced with the data we now possessâthat life appeared as soon as environmental conditions permitted such an event and then remained pervasive forever afterâour thoughts must move to ideas about almost predictive inevitability. Given a planet of earthly size, distance, and composition, life of simplest grade probably originates with virtual certainty as a consequence of principles of organic chemistry and the physics of self-organizing systems.
But whatever the predictability of life's origin, subsequent pathways of evolution have run in mighty peculiar directions, at least with respect to our conventional hopes and biases. The broadest pattern might seem to confirm our usual view of generally increasing complexity, leading sensibly to human consciousness: after all, the early earth sported only bacteria, while our planet now boasts people, ant colonies, and oak trees. Fair enough, but closer scrutiny of general timings or particular details can promote little faith in any steady pattern. If greater size and complexity provide such Darwinian blessings, why did life take so long to proceed “onward,” and why do most of the supposed steps occur so quirkily and so quickly? Consider the following epitome of major events:
Fossils, as stated above, appear as soon as they possibly could in the geological record. But life then remains exclusively at this simplest so-called prokaryotic grade (unicells without any internal organelles, that is, no nuclei, chromosomes, mitochondria, and so on) for about half its subsequent history. The first unicells of the more complex eukaryotic grade (with the conventional organelles of our high school textbook pictures of an amoeba or paramecium) do not appear in the fossil record until about two billion years ago. The three great multicellular kingdoms of plants, fungi, and animals arise sub-sequently (and, at least for algae within the plant kingdom, more than once and independently) from eukaryotic unicells. Fossils of simple multicellular
algae extend back fairly reliably to about one billion years, and far more conjecturally to as many as 1.8 billion years.
But the real enigmaâat least with respect to our parochial concerns about the progressive inevitability of our own lineageâsurrounds the origin and early history of animals. If life had always been hankering to reach a pinnacle of expression as the animal kingdom, then organic history seemed in no hurry to initiate this ultimate phase. About five-sixths of life's time had elapsed before animals made their first appearance in the fossil record some 600 million years ago. Moreover, the earth's first community of animals, which held nearly exclusive sway from an initial appearance some 600 million years ago right to the dawn of the Cambrian period 543 million years ago, consisted of enigmatic species with no clear relation to modern forms.
These so-called Ediacaran animals (named for the locality of first discovery in Australia, but now known from all continents) could grow quite large (up to a few feet in length), but apparently contained neither complex internal organs, nor even any recognizable body openings for mouth or anus. Many Ediacaran creatures were flattened, pancakelike forms (in a variety of shapes and sizes), built of numerous tubelike sections, complexly quilted together into a single structure. Theories about the affinities of Ediacaran organisms span the full gamutâfrom viewing them, most conventionally, as simple ancestors for several modern phyla; to interpreting them, most radically, as an entirely separate (and ultimately failed) experiment in multicellular animal life. An intermediate position, now gaining favorâa situation that should lead to no predictions about the ultimate outcome of this complex debateâtreats Ediacaran animals as a bountiful expression of the range of possibilities for diploblastic animals (built of two body layers), a group now so reduced in diversity (and subsisting only as corals, jellyfishes, and their allies) that living representatives provide little understanding of full potentials.
Modern animalsâexcept for sponges, corals, and a few other minor groupsâare all triploblastic, or composed of three body layers: an ectoderm, forming nervous tissue and other organs; mesoderm, forming reproductive structures and other parts; and endoderm, building the gut and other internal organs. (If you learned a conventional list of phyla back in high school biology, all groups from the flatworms on “up,” including all the “big” phyla of annelids, arthropods, mollusks, echinoderms, and vertebrates, are triploblasts.) This three-layered organization seems to act as a prerequisite for the formation of conventional, complex, mobile, bilaterally symmetrical organisms with body cavities, appendages, sensory organs, and all the other accoutrements that set our
standard picture of a “proper” animal. Thus, in our aimlessly parochial manner (and ignoring such truly important groups as corals and sponges), we tend to equate the problem of the beginning of modern animals with the origin of triploblasts. If the Ediacaran animals are all (or mostly) diploblasts, or even more genealogically divergent from triploblast animals, then this first fauna does not resolve the problem of the origin of animals (in our conventionally limited sense of modern triploblasts).
The story of modern animals then becomes even more curious. The inception of the Cambrian period, 543 million years ago, marks the extinction, perhaps quite rapidly, of the Ediacara fauna, and the beginning of a rich record for animals with calcareous skeletons easily preserved as fossils. But the first phase of the Cambrian, called Manakayan and lasting from 543 to 530 million years ago, primarily features a confusing set of spines, plates, and other bits and pieces called (even in our technical literature) the SSF, or “small shelly fossils” (presumably the disarticulated fragments of skeletons that had not yet evolved to large, discrete units covering the entire organism).
The next two phases of the Cambrian (called Tommotian and Atdabanian, and ranging from 530 to about 520 million years ago) mark the strangest, most important, and most intriguing of all episodes in the fossil record of animalsâ the short interval known as the “Cambrian explosion,” and featuring the first fossils of all animal phyla with skeletons subject to easy preservation in the fossil record. (A single exception, a group of colonial marine organisms called the
Bryozoa, make their appearance at the beginning of the next or Ordovician period. Many intriguing inventions, including human consciousness and the dance language of bees, have arisen since then, but no new phyla, or animals of starkly divergent anatomical design.)

Time charts for major events in the history of life (left) and for details of the Cambrian Explosion and other events in the origin of multicellular animals (right)
.
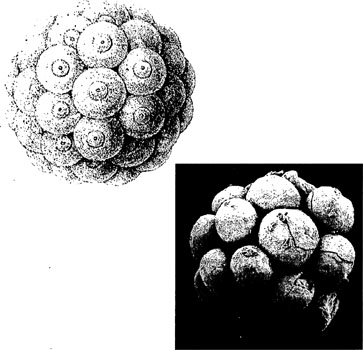
Haeckel's theoretical drawings of ancestral animals (left) compared with fossils of a Precambrian embryo (below)
.
The Cambrian explosion ranks as such a definitive episode in the history of animals that we cannot possibly grasp the basic tale of our own kingdom until we achieve better resolution for both the antecedents and the unfolding of this cardinal geological moment. A major discovery, announced in February 1998, and also based on learning to look in a previously unsuspected place, has thrilled the entire paleontological community for its promise in unraveling the previously unknown history of triploblast animals before the Cambrian explosion.
If the Cambrian explosion inspires frustration for its plethora of dataâtoo much, too confusing, and too fastâthe Precambrian history of triploblast animals engenders even more chagrin for its dearth. The complex animals of the
explosion, so clearly assignable to modern phyla, obviously didn't arise
ex nihilo
at their first moment of fossilization, but who (and where) are their antecedents in Precambrian times? What were the forebears of modern animals doing for 50 million prior years, when Ediacaran diploblasts (or stranger creatures) ruled the animal world?