Life on a Young Planet (37 page)
Read Life on a Young Planet Online
Authors: Andrew H. Knoll

However, there is a problem. At the beginning of this chapter, I stated that the late Proterozoic Earth experienced at least four ice ages, two of global dimensions. But Ediacaran animals and diverse algae follow only the last ice age—earlier Proterozoic glaciation did not engender recognizable evolutionary innovations. Ice ages were not magic wands of evolution.
We have to ask what was different about the aftermaths of younger and older ice ages. I believe that oxygen made all the difference.
In 1959, J. R. Nursall, a zoologist at the University of Alberta, proposed that animals appeared so late in the evolutionary day because only in the latest Proterozoic did our atmosphere accumulate enough oxygen to sustain metazoan physiology. Nursall based his hypothesis strictly on comparative biology—I’m not sure he ever knowingly looked at a Proterozoic rock—nonetheless, his proposal has enjoyed episodic popularity among paleontologists. Only in the past decade, however, has Nursall’s idea garnered empirical support from geology.
There is no question about animals’ need for oxygen. In modern marine basins, the abundance and diversity of animal species declines precipitately as we approach anoxic waters. Small animals persist better than large—nematodes and other tiny animals that inhabit the spaces between sand grains on the seafloor have especially low oxygen needs. Heavily skeletonized species fare most poorly; few armored animals live in oxygen-starved environments.
As early as 1919, August Krogh showed that when marine animals depend on diffusion to supply their tissues with oxygen, body size is limited by the amount of oxygen in surrounding waters. Krogh’s biophysical rule can be circumvented in many ways—animals like
cnidarians (and, probably, the vendobiontids in Ediacaran assemblages) achieve large size by draping thin layers of metabolically active tissue around inert “jelly” or fluid. Circulation of body fluids and specialized respiratory organs (gills or lungs) help as well, carrying oxygen efficiently to distant tissues. Nonetheless, on the Proterozoic Earth, before animals evolved sophisticated circulatory systems, oxygen levels must have determined the effective sizes of animals.
3
The application to late Proterozoic biological history is obvious. Microscopic animals with scant oxygen requirements could have plied Proterozoic seas long before the Ediacaran epoch. Only with a latest Proterozoic rise in oxygen, however, did macroscopic (and, hence, easily fossilizable) animals become possible.
The oxygen hypothesis reconciles paleontology and molecular clocks, and it explains the simultaneous radiations of animals and algae in latest Proterozoic oceans—recall from
chapter 9
that only in oxygen-rich oceans would large-celled eukaryotic algae have gained prominence across continental shelves. As outlined above, a latest Proterozoic rise in oxygen also makes Schrag and Hoffman’s proposal for methane-driven glacial initiation possible and, if that model is correct, explains why global ice ages never returned in the age of animals.
Did any such event leave its mark in the geochemical record? The answer is yes. When John Hayes, Jay Kaufman, another lab alumnus, and I first documented high C-isotopic values in late Proterozoic carbonates, we recognized this as a smoking gun for oxygen influx. As discussed in
chapter 6
, photosynthesizing cyanobacteria and algae use carbon dioxide and water to produce both organic molecules and oxygen. Respiring organisms react the organic matter and oxygen back to carbon dioxide and water. So, as long as photosynthesis and respiration remain in balance, the environment doesn’t change. Burial of organic matter breaks this metabolic couple, making it possible for oxygen to accumulate in the atmosphere and oceans. High C-isotopic values imply that in late Proterozoic oceans, organic matter was buried at unusually high rates. Indeed, when Lou Derry, Jay Kaufman, and Stein Jacobsen used
C-isotopic and other geochemical data to model late Proterozoic atmospheric change, they concluded that oxygen increased strongly just after the younger of the global ice ages—supplying an answer to the question of why life responded differently in the aftermaths of earlier and later glaciations on the late Proterozoic Earth.
Independent evidence for latest Proterozoic oxygen increase comes from the sulfur isotopic record. In
chapter 6
, I outlined the thinking that led Don Canfield to propose that mid-Proterozoic oceans featured moderate oxygen at the surface and hydrogen sulfide at depth. One of Don’s key observations was that S-isotopic fractionation didn’t reach modern values until the end of the eon. The sulfidic ocean returns here—now, however, seen in its closing moments. Only as the Proterozoic Eon ended did sulfur-bearing minerals begin to record the fractionation expected in oceans full of oxygen. Sulfur isotopes also suggest that sulfate concentrations rose to near modern levels at this time, consistent with an overall increase in Earth’s surface oxidation level.
Thus, geochemistry increasingly supports Nursall’s hunch that animal evolution was stirred by oxygen. And with more oxygen, a new world began to emerge. Seaweeds and planktonic algae diversified across continental shelves. Among animals, developmental mutations favoring large body size stopped being lethal and began to be advantageous, introducing new functional possibilities. By 555 million years ago, large size had evolved in colonial protozoans, sponges, cnidarians (and vendobionts), and stem bilaterians.
Perhaps there is nothing more to explain. Perhaps, with the genetic tool kit in place, removal of the oxygen barrier simply allowed animal life to unfold. But there is one more event to explore.
Namibian fossils demonstrate that Ediacaran organisms reigned over marine communities until the very end of the Proterozoic Eon. Conversely, the Kotuikan cliffs show that bilaterian diversity (literally) took shape later, as the Cambrian began. Intriguingly, at the boundary between these two faunas, C-isotopic values fall through the floor, signaling a large, but short-lived, perturbation in Earth’s carbon cycle.
Around the world, carbonate rocks near the Proterozoic-Cambrian boundary display C-isotopic values as low as or lower than those in the cap carbonates that cover late Proterozoic glacial beds (
figure 11.6
).
However, there is little evidence of tillites at the boundary, nor do we find any of the unusual sedimentary features that mark postglacial caps. Radiometric dates tell us that the isotopic anomaly came and went in less than a million years. And research on trace elements (especially uranium enrichment) by Japanese geochemists Hiroto Kimura and Yoshio Watanabe suggests that coastal oceans were transiently starved of oxygen.
What could have caused such a perturbation? We don’t know for sure, but the candidate list is short, and all plausible explanations imply bad times for biology. Moreover, similar chemistry marks a major event in more recent Earth history: the great mass extinction at the boundary between the Permian and Triassic periods. Permo-Triassic catastrophe has been blamed on great bursts of methane from icy stores on continental slopes or catastrophic overturn of the hemisphere-scale Panthallassic Ocean, bringing oxygen-poor but CO
2
-rich waters to the surface. Impact by a meteor or comet also attracts interest, although evidence for extraterrestrial influence on end-Permian extinction remains controversial. The same suspects form the lineup for end-Proterozoic environmental perturbation, but, as in the Permian, we don’t yet know which party, if any, is guilty.
Could mass extinction explain the stratigraphic break and morphological gulf between Ediacaran and Cambrian animals? I think that it can.
4
In fact, marine life across the Proterozoic-Cambrian boundary reminds me a great deal of land animals at another time of mass extinction, the Cretaceous-Tertiary boundary. Beginning near the end of the Triassic Period, dinosaurs dominated terrestrial ecosystems for nearly 150 million years. Mammals shared the landscape for pretty much all of this interval, but remained small and simple. Then, at the Cretaceous-Tertiary boundary, 65 million years ago, a giant meteor slammed into our planet, eliminating the dinosaurs (save for the ecologically distinct birds) but not all mammals. In the ecologically permissive world that followed, mammalian survivors diversified rapidly, giving rise to the
major groups of mammals that have graced plains and forests ever since. I like the idea that Ediacaran animals were the “dinosaurs” of late Proterozoic ecosystems, simple but ecologically effective organisms that held stem-group bilaterians in check. When Ediacarans vanished, they left a huge opening for bilaterian survivors. Bilaterian animals radiated in this relatively empty world, filling it with the fauna that still dominates the oceans—permissive ecology coming into play once more (
figure 12.3
).
This argument has been criticized as hard to test. Yes, Ediacaran diversity plummeted at the Proterozoic-Cambrian boundary,
5
but as increased burrowing began to churn sediments in the Cambrian, the preservational window through which we view Ediacaran biology also closed. This isn’t quite true, but it is true enough to limit our confidence in interpreting the disappearance of Ediacaran fossils from the rock record. (The Burgess Shale and its earlier Cambrian counterparts do afford an opportunity to look for soft-bodied animals. One or two Ediacaran holdovers have been claimed, but by the end of the Early Cambrian, Ediacaran-grade organisms clearly played a minor role at best in marine ecosystems.)
But, there is another record to test. Remember that microbial reefs in Nama and other late Proterozoic successions contain abundant and modestly diverse skeletons of early animals. Working in Oman, where the C-isotopic boundary event can be traced throughout a large and well-explored basin (it hosts a giant oil field!), John Grotzinger found that
Cloudina
,
Namacalathus
, and other skeletal fossils occur abundantly in all reefs up to the horizon of the isotopic excursion. The microbial reefs continue above it, but they don’t contain skeletons. They are almost eerily quiet, much like sedimentary beds just above the Permian-Triassic boundary. Thanks to John, then, we have positive evidence for a biological change of guard precisely coincident with environmental insult at the end of the Proterozoic.
As the Cambrian dawned, there may have been a moment—in a world depleted by extinction and before genetics had locked animals into particular patterns of growth and development—when all things were possible in animal evolution. Any such moment, however, must have been brief. Guided by developmental genetics, expanding animal populations began to accumulate the biological features we associate today with arthropods and brachiopods, echinoderms and chordates. And with emerging body plans came differing functional possibilities that partitioned the metazoan world and shaped the connections among species. Algae diversified, as well, in a Cambrian Explosion that cut across kingdoms.
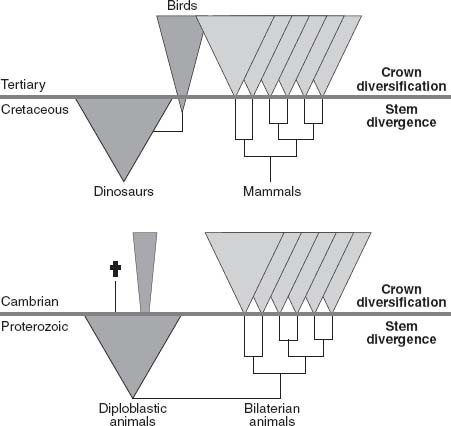
Figure 12.3.
Diagram illustrating the suggested analogy between evolution on land at the Cretaceous-Tertiary boundary and evolution in the oceans at the Proterozoic-Cambrian boundary. Diploblastic animals include sponges, cnidarians, and probably, most Ediacaran organisms. (Reprinted with permission from A. H. Knoll and S. Carroll, 1999. Early animal evolution: emerging views from comparative biology and geology.
Science
284: 2129–2137. Copyright 1999 American Association for the Advancement of Science)
Physical events may thus have provided the opportunity for Cambrian
diversification. But the evolutionary paths actually traveled by Cambrian animals reflect the interplay between development and ecology. Predators and prey locked into an evolutionary arms race, while grazers and algae began to shape the limits of each other’s existence. More than ever before, biological interactions and not just the physical environment determined the shape of life. And as the world filled ecologically, evolutionary opportunities for further new body plans dwindled. In the seas, the hand that animal evolution would play for the next 500 million years had been dealt.
Beneath this new ecological edifice, of course, Earth’s age-old ecological circuitry continued unchanged. As they did 3 billion years earlier, bacteria continued to cycle biologically important elements through ecosystems, sustaining the biosphere that made animal life possible.
Truth in advertising forces me to acknowledge that some paleontologists tell the story of animal diversification without invoking any environmental influence. In this telling, animals arose only in latest Proterozoic oceans, quickly taking evolutionary shape as the Ediacaran fauna—which, in the predatory world of the Cambrian, gave way to more sophisticated crown group bilaterians and cnidarians. Viewed this way, genetics and ecology are not just important drivers of the Cambrian evolution; they are the only drivers.
The idea that late Proterozoic evolution owes nothing to environmental change has some articulate adherents, but I believe that it flies in the face of everything geology has taught us about both the late Precambrian Earth and more recent evolution. If there is one lesson that paleontology offers to evolutionary biology, other than the documentation of biological history itself, it is that life’s opportunities and catastrophes are tied to Earth’s environmental history. We can only understand macroevolution—the comings and goings of species and higher taxa through time—if we link the microevolutionary processes studied by geneticists with Earth’s dynamic environmental history. The great physical events that framed early animal evolution—global glaciation, the rise of oxygen-filled oceans, and extraordinary perturbations of the carbon cycle—are among our planet’s most profound environmental events. We ignore them at our peril.
And here, with the seeds of modern biology planted, my narrative ends. The Cambrian Explosion—both the culmination of life’s long Precambrian history and a radical departure from it—emerges as the product of unique interactions between biological and physical processes. The developmental tool kit built by the duplication, mutation, and rearrangement of genes was necessary for animal diversification, but it likely came together at least 600 million years ago and, by itself, could not complete the biological revolution. The new biology required permissive ecology so that unusual genetic variants could survive, and surviving variants to provide the raw material for morphological innovation. In the great physical upheavals that ended the Proterozoic Eon, genetic possibility and environmental opportunity together spawned new and diverse ecosystems in the world’s oceans.