Junk DNA: A Journey Through the Dark Matter of the Genome (24 page)
Read Junk DNA: A Journey Through the Dark Matter of the Genome Online
Authors: Nessa Carey

In order to understand this process, we have to be a little more sophisticated than usual in the way we think of the genome. Much of the time it’s very helpful to describe DNA in terms of a ladder, or railway lines, because it helps us to visualise the two strands and the way they are held together by base pairs. But the problem with this is it makes us think in very linear terms. We probably also think of DNA as being quite a stiff molecule because subconsciously we are comparing it with solid artefacts from our more familiar physical surroundings.
But we already recognise that DNA isn’t a stiff molecule, because we know that it can be squashed up and compacted really dramatically to fit into the nucleus. So let’s explore that a bit more. If we take the double-stranded nature of DNA as a given (so as not to complicate the picture) we can imagine a section of our genome as being like a very long piece of pasta, maybe the longest piece of tagliatelle ever created. This is marked in a couple of places by food dye, representing the enhancer and the protein-coding gene. Looking at Figure 12.3 we can see two scenarios. When the pasta is uncooked, it’s very inflexible and the enhancer and gene are far apart. But if the pasta is cooked, the tagliatelle becomes flexible. It can fold and bend in all sorts of directions and these can bring the dyed regions representing the enhancer and gene together.
Some parts of our chromosomes are repressed and shut down almost permanently in different cells, to switch off genes that never need to be expressed in that tissue type. Our skin cells, for example, don’t need to express the proteins that are used to carry oxygen around in the blood. These genomic regions are completely inaccessible in the skin cells, curled up tight like an over-wound spring. But there are huge regions that aren’t in this hyper-condensed state and where genes are accessible and can potentially be switched on. In these sections the DNA is like the cooked pasta, like having the
longest piece of tagliatelle in the world, filling an entire pot on its own. It bends and swirls in the cooking water, throwing out loops and arcs.
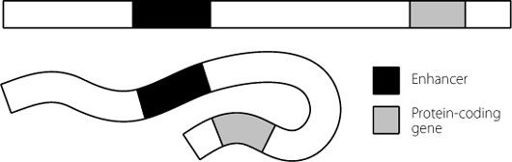
Figure 12.3
Simple schematic to show how folding of a flexible DNA molecule can bring two distant regions, such as an enhancer and a protein-coding gene, into close proximity to each other.
In this way, a protein-coding gene and its distant enhancer may come very close to each other. The long non-coding RNA and the Mediator complex then hold the two loops together and ensure that expression of the gene is driven up. Another complex also has to work with the Mediator complex to carry this out.
e
The additional complex is one that’s also required for separating duplicated chromosomes during cell division, so it’s well equipped for dealing with large-scale movements of DNA. Mutations in some of the genes that encode members of this additional complex cause two developmental disorders, called Roberts Syndrome and Cornelia de Lange syndrome.
17
The precise features of the affected children can be quite variable, probably depending on the exact gene that is mutated, and the precise mutation in that gene. Typically, the children are born small and remain relatively undersized; they have a learning disability and frequently present with limb abnormalities.
18
The extent of this looping mechanism is quite remarkable, and may not just be restricted to enhancers. It may also be used to bring other regulatory elements close to genes. In a study of three human cell types, analysing just 1 per cent of the human genome, researchers identified over 1,000 of these long-range interactions in each cell line. The interactions were complex, most frequently involving regions that were separated by about 120,000 base pairs. Often the regulatory region looped up to a gene that wasn’t the nearest one to it. In fact, in over 90 per cent of the loops, the nearest gene had been ignored. Think of this as needing to borrow a cup of sugar and visiting someone half a mile away instead of popping in to your next door neighbour.
And if we continue the neighbour theme, the relationships were outrageously promiscuous. Imagine a 1970s partner-swapping party on steroids. Some genes interacted with up to twenty different regulatory regions. Some regulatory regions interacted with up to ten different genes. These probably don’t all occur in the same cell at the same time. But what they show is that there is not a simple A to B relationship between genes and regulatory regions. Instead, there is a complex net of interactions, giving a cell or an organism an extraordinary amount of flexibility in how it regulates its overall tapestry of gene expression.
19
Although there is still plenty to be unravelled about the networks and how they operate, it would appear that while the junk DNA that forms promoters switches on our genomic engines, it’s the junk DNA that forms long non-coding RNAs and enhancers that converts that engine from one powering a Sandero to one that can accelerate the Veyron down the freeway of life.
From cottage industry to the factory floor
Remarkable though the looping between individual regulatory regions and genes undoubtedly is, there is an even more dramatic set of long-range interactions that occur in cells. To understand the significance of this, a short social history lesson may be in order. In the early part of the 19th century in Britain, the bulk of all textile work was carried out as a cottage industry. Essentially, individuals worked in their homes on small-scale production. If you had mapped out the locations of textile production in a given region, you’d have a map with lots of individual dots on it, where each working cottage was located. Fast-forward about 50 years and into the Industrial Revolution and the same study would create a very different picture. Instead of a fairly homogenous dotted distribution, like a pointillist painting, you’d find a map with just a few large spots showing the location of big factories.
Even if we just think about the protein-coding genes, we know that thousands are typically switched on in a given human cell type. These genes are spread out across our 46 chromosomes, so we might expect that if we analysed a cell to visualise the geographical locations of the genes that are switched on we would see thousands of tiny dots spread throughout the nucleus. Instead, as shown schematically in Figure 12.4, there are about 300 to 500 larger spots.
20
Gene expression in our cells isn’t a cottage industry. Instead it takes place in discrete locations in the nucleus known as factories.
21
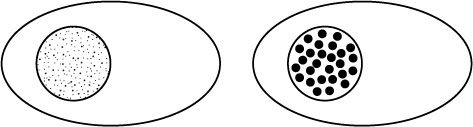
Figure 12.4
The dots represent the positions of protein-coding genes in the nucleus. If genes were positioned in the nucleus solely as a function of their position on chromosomes, we would see a diffuse pattern such as the one on the left. Instead, genes cluster together in three-dimensional space, creating a punctate pattern of gene localisation represented by the situation on the right.
Each factory contains between four and 30 copies of the enzyme that makes a messenger RNA molecule from the DNA template, plus a large number of other molecules required to do the work.
22
,
23
The enzymes stay in one place and the relevant gene is reeled through to be copied.
24
In order for the gene to reach the factory, the DNA has to loop out to reach the right part of the cell nucleus. But the really ingenious bit is that more than one gene can be copied into messenger RNA at a time in the same factory. The combination of genes found in a single factory isn’t random. The genes tend to be ones that code for proteins that are used for related functions in the cell. This is equivalent to having multiple
parallel assembly lines in one physical factory. Once all the lines have completed their individual tasks, the factory can assemble the final product from the components. One factory produces boats, another builds food mixers. In our cells, the factories ensure that genes are expressed in a coordinated fashion. This means lots of loops unfurling from chromosomes and localising to the same regions of the nucleus simultaneously.
One example of this is a factory for the genes that code for the proteins required to create the complex haemoglobin molecule, which carries oxygen around in the blood.
25
Another factory is used to generate the proteins required in order to mount a strong immune response.
26
One important component of an effective immune response is the production of proteins called antibodies. Antibodies circulate in the blood and other body fluids, binding to any foreign matter that they detect. Scientists activated the cells that produce antibodies and then studied how certain key genes looped out. The genes they analysed were the ones required to create antibody molecules. They found that these key genes moved to the same factory as each other. Remarkably, some of these genes were completely physically separate from each other normally, as they are located on different chromosomes.
Although this is a remarkable way of coordinating gene expression, it may also carry risks. Burkitt’s lymphoma is the aggressive cancer we met earlier in this chapter. The cell type that becomes abnormal in this disease is the cell type that produces antibodies. In this condition, a strong promoter from one chromosome gets abnormally positioned next to a gene from another chromosome. Until recently we didn’t understand why these regions were susceptible to joining up, because we thought of them as being physically distant from each other, as they are on different chromosomes. But now we know that the regions that ‘swap’ to create the dangerous abnormal hybrid chromosome are both regions that move to the factory described in the previous paragraph.
This might be how the two different chromosomes get close enough together to swap their material, perhaps if both break simultaneously and are wrongly repaired when in the factory.
While it might seem that evolution would have selected against this dangerous situation, we need to remember yet again that natural selection is about compromise, not perfection. The advantages of producing antibodies to fight off infections and thereby keep us alive long enough to reproduce clearly outweigh the potential disadvantages of an increased cancer risk.
Footnotes
a
The gene codes for a protein called MYC. MYC is also involved in a range of other cancers.
b
The modification is addition of a phosphate group (one phosphorus atom and four oxygen atoms) to a specific position on histone H3. This modification is usually associated with active genes.
c
The Mediator component is MED12.
d
These are known as induced pluripotent stem cells (iPS cells).
e
This additional complex is called Cohesin.
13. No Man’s Land
When we think about the First World War, the prevailing image many of us have is probably of men in trenches. Opposing armies dug into the muddy landscapes for the ultimate exposition of war as months of boredom, punctuated by moments of acute terror.
1
The trenches occupied by the armies were separated by stretches of terrain that were not under the control of either combatant. These stretches were ‘No Man’s Land’, and could be as narrow as a couple of hundred metres, or over a kilometre wide. At night, the soldiers would creep out of the trenches for reconnaissance, to lay barbed wire and to retrieve injured or dead compatriots.
The human genome contains multiple regions of No Man’s Land, keeping different elements apart from each other. Just like the quagmires of the First World War, these genomic barriers vary in size and are fairly fluid, depending on where they lie in relation to their troop movements. And just like the No Man’s Land of Europe in those awful few years of slaughter, these regions are anything but devoid of activity. The No Man’s Land of the human genome binds proteins, garners epigenetic modifications and regulates the interactions of different genetic elements in a highly active way.