The Puzzle of Left-Handedness (26 page)

Over time a number of facts have emerged that give us something solid to grasp in our search for the causes of hand preference (and indeed for its ultimate consequences, but more of that later).
One such firm fact is that although the results of research are never exactly the same when tests are repeated, many studies large and small show that left-handedness is slightly more common in men than in women. Ten as opposed to 9 per cent; 14 as opposed to 12 per cent: those are the kinds of figures produced.
We can also be fairly certain that people with brain damage, or people in whom brain damage is suspected, no matter how slight, are rather more likely than average to be left-handed. Based on this finding, scientists have tried to discover whether there is a connection between left-handedness and problematic births to older mothers, and these tests quite often show slightly elevated numbers. All in all, we cannot entirely dismiss the notion that left-handedness may sometimes be the result of damage to the brain.
Brain damage certainly cannot explain all cases of left-handedness, however. For that there are simply too many right-handers with evidence of brain damage and far too many left-handers in whom no abnormalities can be found. We must therefore assume that left-handedness can also arise in some other way.
Another fact that emerges repeatedly is that the percentage of left-handers in the population falls sharply with age. Variations on the saying ‘anyone who isn’t on the left at twenty has no heart; anyone who’s still on the left at forty has no brain’ have been attributed to a range of famous men, from Winston Churchill and Georges Clemenceau to Aristide Briand, but always with reference to politics. In some bizarre way it seems to apply to hand preference as well, since between the ages of twenty and forty the proportion of the population that calls itself left-handed declines significantly.
There is also firm evidence for an inherited component, one indication being the demonstrable connection between hand preference in parents and children. Couples in which one partner is left-handed are roughly twice as likely to have a left-handed child. (With two right-handed parents the chances are about 10 per cent.) If both parents are left-handed, the figure doubles again, approaching 40 per cent. We might initially think this could be a product of imitation, of setting an example that is then followed, but that’s extremely unlikely. Firstly because the majority of left-handers have two right-handed parents and the majority of children of left-handed parents are right-handed, and secondly because there are indications that in at least some cases hand preference develops before birth. Around 5 per cent of unborn babies suck their left thumbs in the womb, and this seems later to translate into left-handedness. Parents couldn’t possibly have any influence on this kind of behaviour in an unborn child.
Another indication that inherited factors are at work is that left-handedness occurs roughly twice as often in monozygotic twins as it does in the products of single births. A thoroughly mystifying fact is that the same goes for dizygotic twins.
It’s therefore clear that we need to focus our search for the causes of hand preference on hereditary influences and minor mishaps. These are the basis for the two most important theories doing the rounds today, the genetic theory put forward by Marian Annett and the hormonal testosterone theory propounded by Norman Geschwind.
29
Genetic Left-handedness
In the twentieth century several attempts were made to explain left-handedness by reference to genetic differences. Most came to nothing. This was mainly because the distribution of left- and right-handedness did not coincide with the laws of inheritance developed by Gregor Mendel, a shy Austrian monk who by studying the peas in his garden unravelled the fundamentals of how hereditary characteristics are passed on from generation to generation and the patterns according to which they manifest themselves.
Mendel made his discoveries based on the external appearance of pea plants. He knew almost nothing about what was going on inside them. We now know that every ordinary cell of a plant, animal or human contains inheritable material, almost all of it in the cell nucleus. It’s organized into chromosomes, tiny coils of immensely long spaghetti-like strings of protein and
DNA
, pieces of which, known as genes, contain a recipe for making one or other protein, or for controlling the activities of other pieces of
DNA
. Together the genes of all the chromosomes in an organism form what has been described as a book containing the instructions for making it. A living organism is by far the most complicated thing in the world, so it’s a monumental work, sometimes hundreds of volumes long.
In the case of human beings there are 23 volumes. In other words, the nucleus of each of the cells in our bodies contains 23 different chromosomes. Since they all occur in pairs, the nucleus of every normal human cell contains a total of 46, and in every case except one, each pair consists of two copies of the same chromosome.
The 23rd pair is the exception, since only in women are the two chromosomes the same. That’s what makes a woman a woman; we are talking here about the sex chromosomes x and y. Women have two copies of the
X
chromosome, whereas in men one copy of the
X
chromosome is paired with one copy of the
Y
chromosome. The possession of a
Y
chromosome is what it means to be male, from a biological perspective. It’s responsible for the inheritable characteristics that make a man different from a woman.
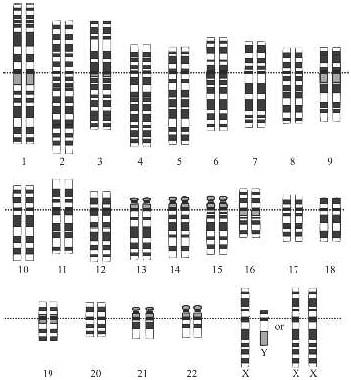
Stylized representation of the chromosomes in the nucleus of a human cell. The ordinary chromosomes, known as autosomes, are numbered from the largest to the smallest. The sex chromosomes are called
X
(female) and
Y
(male). At the bottom right are the two possible pairs. A man has the pair
XY
, a woman the pair
XX
.
Were it the case that at conception two normal cells, one from the mother and one from the father, simply merged, then the resulting child would have not two but four copies of each chromosome. The next generation would have eight, its children sixteen and so on until in a relatively short time the cell walls would literally burst under the pressure of billions of chromosomes. Instead, reproduction is exclusively the task of special reproductive cells: a woman’s eggs and a man’s sperm. At the point when they are ripe and ready to do their work, they contain only one copy of each chromosome. When an egg cell merges with a sperm cell, the resulting cell has two copies of each chromosome, sometimes including an
XY
pair, sometimes an xx pair.
A child therefore inherits one complete set of chromosomes from its father and one from its mother. Still, it’s plain to see that children are never simply a half-and-half mix of their parents. There are two main reasons for this.
The first is the existence of gene variants. Many genes occur in different forms, with different effects. Sometimes the results are slight variations, such as differences in hair colour, or in the colour of the skin and eyes. Sometimes certain variants cause illness; they are the source of genetic disease. The variants of a gene, also known as alleles, are subject to rules of precedence. Some, described as recessive, will be pushed aside if the chromosome with which they are paired contains a different variant, taking effect only if there are two of them. Other alleles are dominant, in other words: what they say goes. Since a father and mother each contribute one copy from each of their pairs of genes to their child’s chromosomes, combinations of characteristics will occur in the child that are not seen in either of its parents. So although children resemble their parents, they are not born with a straightforward combination or a wholly representative sample of their parents’ characteristics.
The second cause of variation between children with the same father and mother is a phenomenon known as recombination. Evolution is a knock-out race, governed by chance, and it has rendered up a system of reproduction that guarantees as much variation as possible. This gives a species the best chance of survival, since where a multitude of types is created, there are always one or two that can flourish in the circumstances that happen to prevail at any given time. Recombination means that during the process that will eventually produce reproductive cells with one copy of each chromosome, the two copies are first thoroughly whisked together. The single chromosome that is passed on is therefore not exactly the same as either of the copies in the cells of the parent but rather a random mix of the two. This ensures that children don’t exactly resemble their parents in every respect, and it also means they are a bit like just about all their forebears. The shuffling of cards that we call recombination means that from each of a person’s many ancestors, one or two characteristics are likely to show up many generations later. Recombination is also responsible for the fact that aside from gender, no one precisely resembles a brother or sister. It makes the genetic constellation in each egg and each sperm unique.
Mendel’s laws describe the rules of precedence that apply to gene variants and the results they produce, although only in the case of characteristics that arise from a single gene. Hair colour is one such characteristic. If we assume for the sake of convenience that just two colours exist, fair and dark, then there’s a gene for hair colour that has two variants. Let’s call them
F
for fair hair and
D
for dark hair. We all inherit either an
F
or a
D
allele from each of our parents.
If a baby boy inherits a D allele from both his father and his mother, then he is guaranteed to have dark hair. After all, there is nothing in his genes that could cause blondness. For the same reason, everyone with two
F
variants is fair-haired. But the assumption that children who receive an
F
allele from one parent but a
D
from the other will be dark blonde is wrong. They will all have dark hair, because the
D
allele is dominant. If a
D
variant coincides with an
F
variant, the
D
variant takes precedence. So the
F
variant is recessive.
Anyone with fair hair must therefore have the combination
F-F
, while anyone with dark hair may have inherited either
D-D
or
D-F
. This has consequences of its own. Although two fair-haired people will only ever have fair-haired children, since between them they have no
D
variant to pass on, two dark-haired people can have children some of whom are dark and others fair. If both parents are
D-F
and both pass on an
F
allele, the result is
F-F
and therefore fair-haired. Recessive alleles can therefore be handed down unnoticed for several generations before suddenly being expressed.
This may seem rather capricious, but the overall proportion of people with characteristics that are determined by single genes is in theory predictable. If both alleles are equally common, a quarter of the population will have two copies of the recessive variant, a quarter will have two copies of the dominant allele, and the rest will have one dominant and one recessive copy. This means that three-quarters of the population will display the dominant characteristic.
The occurrence of left-handedness cannot be explained based purely on the distinction between dominant and recessive alleles, since the number of left-handed people comes nowhere close to the 25 per cent that would be expected if a recessive allele were responsible. Still, variants do not always occur in equal numbers in a given population. In Scandinavia, for example, fair hair is so common that the F allele of the hair colour gene must be present at an extremely high rate in the population. Outside Europe, and leaving aside communities of people whose forebears emigrated from Europe, you have to search long and hard to find fair hair, since the f allele is extremely rare. Blonde Chinese people exist but there are very few of them indeed. So if the left-handedness allele is relatively uncommon, wouldn’t the actual proportions of left- and right-handers automatically roll out of the mix?
This sounds like a neat solution, but in fact it is a red herring. Differences in frequency between gene variants are the cause of what we call racial characteristics: skin colour, hair colour, the shape and colour of the eyes, and other things of that sort. Even slight isolation in combination with different climatic conditions is sufficient to produce striking superficial differences between peoples over a period of roughly 10,000 years. And they are not confined to useful but otherwise innocent adjustments to features such as eye shape and hair colour. Sickle cell anaemia is a serious, recessive disorder that causes a defect in red blood corpuscles, making them sickle-shaped and unable to do their normal work properly. It’s rare almost everywhere, but in areas where malaria is widespread the incidence of the condition is far higher than normal. The reason is that it offers protection against the even more damaging malaria parasite, which spends part of its life cycle in healthy red blood corpuscles and multiplies far more slowly in sickle cells.
The distribution of hand preference bears no resemblance to any of this. In all times and all places it’s almost precisely the same. Hand preference is therefore a characteristic unaffected by circumstances; there seem to be no selective pressures. Yet sensitivity to selection is at the heart of Mendel’s laws. He played God by crossing different strains and dictating the circumstances under which his plants were allowed to reproduce. The power available to us by this route is demonstrated by the simple fact that the Great Dane, the poodle, the beagle, the Pekinese and the naked chihuahua were all produced in the space of a few thousand years by selective breeding from the same ancestor. We have achieved astonishing variation – yet paw preference in animals has proven impossible to influence by selective breeding.
In a rather desperate attempt to rescue the idea that hand preference follows the pattern of Mendelian inheritance, it has been suggested that the recessive left-handedness allele of a presumed hand-preference gene works badly, so even people with two recessive alleles for left-handedness can end up right-handed. But this is thoroughly unconvincing. For one thing such a gene variant would have to work very badly indeed, given that left-handedness is more common than average in monozygotic twins but in the vast majority of cases affects only one of them. At the same time it must work well enough to continue to produce left-handers, as a hard and fast rule, at a rate of 10 per cent in any given human population.
There is one fact that definitively closes the door on a Mendelian account of the distribution of hand preference. Any explanation in terms of a recessive allele would inevitably mean that two left-handed parents could produce only children who were, genetically at least, left-handed, whereas in fact the majority of children of such parents are right-handed.
The best attempt yet to link left-handedness with genes was made around 1970 by Marian Annett, a psychologist at the University of Hull. It occurred to her that the distribution of hand preference in humans and paw preference in animals, although different, is patently similar. In animals the proportions suggest that paw preference emerges entirely by chance: one quarter are left-pawed, one quarter right-pawed and the rest undecided. The same groups exist in the human population, Annette argued, as long as you ignore those cases of left-handedness that are attributable to some kind of physical damage. Annett believed there was a group of left-handers amounting to around 4 per cent of the population, a group of right-handers that made up some two-thirds of the total and a group of mixed-handed people that accounted for the remaining 30 per cent. If you convert these proportions into a graph, then as with animals you get the bell-shaped curve that statisticians regard as a normal distribution, the difference being that the human curve has been budged a considerable distance towards the right. In other words, by far the larger proportion of the curve – and therefore of people – falls into the right-handed region, while most animals are around the neutral middle point and therefore somewhere in between left-pawed and right-pawed. These are animals that don’t care which paw they use.