The Flamingo’s Smile (16 page)
Read The Flamingo’s Smile Online
Authors: Stephen Jay Gould

(I must stop at this point, for I suddenly realize that I have almost broken my first rule. Scientists have a terrible tendency to present their work as a logical package, as if they thought everything out in careful and rigorous planning beforehand and then merely proceeded according to their good designs. It never works that way, if only because anyone who can think and see makes unanticipated discoveries and must fundamentally alter any preconceived strategy. Also, people get into problems for the damnedest of peculiar and accidental reasons. Projects grow like organisms, with serendipity and supple adjustment, not like the foreordained steps of a high school proof in plane geometry. Let me confess. I was first drawn to
Cerion
because I wanted to compare its fossils with snails I had studied on Bermuda. I studiously avoided all modern
Cerions
because I was petrified by the thicket of available names and considered them intractable. Woodruff first went to Inagua because he wanted to study color banding in another genus of snails. But he went at the height of mosquito season and lasted two days. We took our first trip together to Grand Bahama Island: I to study fossils, he to try the other genus again. But I soon discovered that Grand Bahama has no (or very few) rocks of terrestrial origin, hence no fossil land snails. The other genus wasn’t much more common. We were stuck there for a week. So we studied the living
Cerions
and found a pattern behind the plethora of names. Since then, following Satchel Paige’s advice, we have never looked back.)
About fifteen names had been proposed for the
Cerions
of Grand Bahama and neighboring Abaco Island. After a week, Woodruff and I recognized that only two distinct populations inhabited these islands, each restricted to a definite and different environment.
Abaco and Grand Bahama protrude above a shallow platform called Little Bahama Bank (see accompanying map). When sea level was lower during the last ice age, the entire platform emerged and the islands were connected by land. Little Bahama Bank is separated by deep ocean from the larger Great Bahama Bank, source of the more familiar Bahamian islands (New Providence, with its capital city of Nassau, Bimini, Andros, Eleuthera, Cat, the Exuma chain, and many others). All these islands were also connected during glacial times of low sea level. As Woodruff and I moved from island to island on Great Bahama Bank, we found the same pattern of two different populations, always in the same distinctive environments. On Little Bahama Bank, a dozen invalid names had fallen into this pattern. On Great Bahama Bank, they collapsed, literally by the hundred. About one-third of all
Cerion
“species” (close to 200 in all) turned out to be invalid names based on minor variants within this single pattern. We had reduced a chaos of improper names to a single, ecologically based order. (This reduction applies only to the islands of Little and Great Bahama Bank. Islands on other banks in the southeastern Bahamas, including Long Island, the southeasternmost island of Great Bahama Bank, contain truly different
Cerions
. These
Cerions
can also be reduced to coherent patterns based on few true species. But one essay can treat just so much, and I confine myself here to the northern Bahamas.)
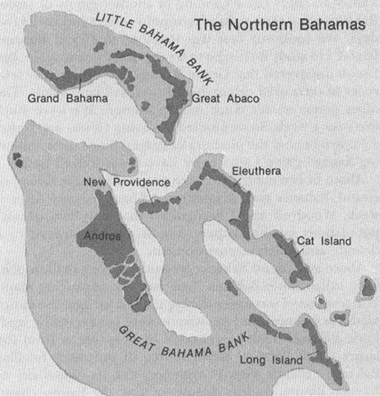
A map of the northern Bahamas, showing edges of the banks.
REPRINTED FROM NATURAL HISTORY
.
Bahamian islands have two different kinds of coastlines. Major islands lie at the edge of their banks. The banks themselves are very shallow across their tops but plunge precipitously into deep ocean at their edges. Thus, bank-edge coasts abut the open ocean and tend to be raw and windy. Dunes build along windy coasts and solidify eventually into rock (often mistakenly called “coral” by tourists). Bank-edge coasts are, therefore, usually rocky as well. By contrast, coastlines that border the interior parts of banks—I will call them bank-interior coasts—are surrounded by calm, shallow waters that extend for miles and do not promote the building of dunes. Bank-interior coasts, therefore, tend to be vegetated, low, and calm.
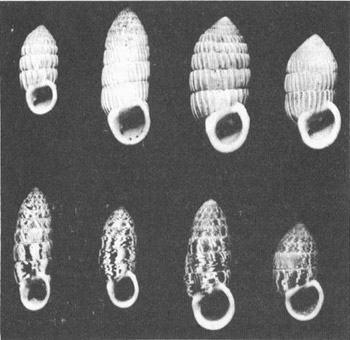
Comparison of ribby, bank-edge
Cerion
(upper row) from Little and Great Bahama Bank, with mottled, bank-interior
Cerion
(lower row).
PHOTO BY AL COLEMAN
.
Woodruff and I found that bank-edge coasts in the northern Bahamas are invariably inhabited by thick-shelled, strongly ribbed, uniformly colored (white to darkish brown), relatively wide, and parallel-sided
Cerions
. To avoid writing most of the rest of this column in Latin, I will skip the formal names and refer to these forms as the “ribby populations” (see photo above). Bank-interior coasts are the home of thin-shelled, ribless or weakly ribbed, variegated (usually with alternating blotches of white and brown), narrow, and barrel-shaped
Cerions
—the “mottled populations.” (Mottled
Cerions
also live away from coasts in the centers of islands, while ribby
Cerions
are confined exclusively to bank-edge coasts.)
This pattern is so consistent and invariable that we can “map” hybrid zones even before we visit an island, simply by looking at a chart of bathymetry. Hybrid zones occur where bank-edge coasts meet bank-interior coasts.
This pattern might seem worthy of little more than an indulgent ho-hum. Perhaps mottled and ribby shells are not very different. Maybe the two environments elicit their differing forms directly from the same basic genetic stock, much as good and plentiful food can make a man fat and paltry fare eventually convert the same gent to a scarecrow. The very precision and predictability of the correlation between form and environment might suggest this biologically uninteresting solution. Two arguments, however, seem to stand conclusively against this interpretation and to indicate that mottled and ribby
Cerions
are different biological entities.
First, the ribby snails are not merely mottled forms with thicker and ribbier shells. As my technical contribution to our joint work, I measure each shell in twenty different ways. This effort permits me to characterize both growth and final adult form in mathematical terms. I have been able to show that the differences between ribby and mottled involve several independently varying determinants of form.
Second, an analysis of hybrid zones proves that they mark a mixture of two different entities, not a smooth blending of populations only superficially separate. My morphological analysis shows, in many cases, the anomalies of form, and the increased variation, that so often occur when two different developmental programs are mixed in offspring. Woodruff’s genetic analysis also proves that the hybrids combine two substantially different systems, for he finds both generally increased genetic variability in hybrid samples, and genes detected in neither parental population.
We can demonstrate that ribby and mottled represent populations with substantial biological differences, but we cannot specify the cause of separation since we have been unable to distinguish between two hypotheses. First, ecological: ribby and mottled forms may be recent and immediate adaptations to their differing local environments. White or light-colored shells are inconspicuous against the bank-edge background of dune rocks, while thick and ribby shells protect their bearers on these windy and rocky coasts. Mottled shells are equally inconspicuous (indeed remarkably camouflaged) when dappled sunlight filters through the vegetation that houses
Cerion
on most bank-interior coasts, while thin and light shells are also well suited for hanging from thin twigs and grass blades. Second, historical: the pattern may be substantially older (although still probably adaptive for the reasons cited above). When sea level was much lower and the banks lay exposed during glacial periods, perhaps ribby populations inhabited all coasts (since all were then bank-edge), while mottled populations evolved for life in island interiors. As sea level rose, ribby and mottled snails simply kept their positions and preferences. The new bank-interior coasts were the interiors of previously larger islands and they continue as homes of mottled snails.
The distinction of mottled and ribby resolved nearly all the two hundred names previously given to
Cerions
from the northern Bahamas. But one problem (involving about ten more names) remained. A third kind of
Cerion
, bearing a thick, but smooth, pure white, and triangularly shaped shell, had been found on Eleuthera and Cat Island. Previous reports indicated nothing about their ecology or habits, but we found these thick white snails in two disjunct areas of southern Eleuthera and in southeastern Cat Island. They prefer island interiors and fit
Cerion
’s general pattern with gratifying predictability—that is, they hybridize with mottled populations as we approach bank-interior coasts and with ribby populations as we move toward bank-edge coasts. But what are they? Just as ecology and genetics resolved the basic pattern of mottled and ribby, we must call upon paleontology to explain our remaining source of diversity.
Fossil dunes of the Bahamas formed at times of high sea level during warmer periods between episodes of glaciation (ice ages). Three major sets of dunes built New Providence, the only Bahamian island with a documented geological pedigree (see Garrett and Gould, in bibliography). These include, from youngest to oldest, a few small dunes less than 10,000 years old and deposited since the last glaciers melted; an extensive set (forming the island’s backbone), representing the high sea levels of 120,000 years ago, before the last glaciers formed; and a smaller set (situated near the island’s center) built more than 200,000 years ago, before a previous glacial period. The oldest dunes contain a fossil
Cerion
now unknown in the Bahamas (see photo on chapter 11). The second and most extensive set includes two species of
Cerion
, a dwarf form now extinct and a large, smooth white species called
Cerion agassizi
(named for Alexander Agassiz, son of Louis, and a pioneer of scientific oceanography in the West Indies). The most recent set, as expected, contains either ribby or mottled
Cerions
, as in the modern fauna. We compared the large white snails of Eleuthera and Cat with
C. agassizi
and found no substantial differences. The small populations on these islands are surviving remnants of a species that once lived in abundance on all the islands of Great Bahama Bank.
The two hundred “species” of northern Bahamian
Cerion
therefore reduce to three basic types with a sensible and ordered distribution. Geographic pattern identified ribby and mottled populations, but we needed an assist from history to understand the smooth white shells of Eleuthera and Cat. It is an awfully long stride from this taxonomic exercise in natural history to our ultimate goal—an understanding of how
Cerion
’s unparalleled diversity of form evolved—but we have taken the first step along the only pathway I know.

Fossils from New Providence Island include (from left to right) an extinct form from the lowest dunes; an extinct dwarf
(Cerion universe)
and the large
Cerion agassizi
from the 120,000-year-old dunes; and a ribby snail from the youngest dunes. Remnant populations of
Cerion agassizi
survive on Eleuthera and Cat Island.
PHOTO BY AL COLEMAN
.