The Cerebellum: Brain for an Implicit Self (38 page)
Read The Cerebellum: Brain for an Implicit Self Online
Authors: Masao Ito
Tags: #Science, #Life Sciences, #Medical, #Biology, #Neurology, #Neuroscience

The final study mentioned here was one in which monkeys were trained to use a rake to obtain food rewards. All three monkeys tested (monkeys E, N, and F) learned to use the rake within the 14-day training period. Monkeys E and F used their right hand to rake and their left hand to retrieve the food reward. Monkey N used both hands to both rake and retrieve the food. They were examined repeatedly in a 4-T scanner for a six-week period: two weeks of habituation to the task, two weeks of intensive daily training, and a two-week post-training period. As task performance improved, structural MRI scans revealed significant increases in the signal intensity of gray matter in the cerebral cortex. These increases were the most significant in the right superior temporal sulcus, right second somatosensory area, and right intraparietal sulcus, with less significant increases in these same regions of the left hemisphere. Interestingly, signal increases were also observed in the white matter of the bilateral cerebellar hemispheres in lobule V (
Quallo et al., 2009
). An observed enhancement of structural MRI signals indicated an increase in the volume of gray and/or white matter. What really underlies the just-described cerebellar enhancement is still unknown, but it is in keeping with long-lasting plasticity in cerebellar circuits that play a role in cerebellar learning.
Neuronal mechanisms for motor actions have been studied by unit recording of Purkinje cell discharge in monkeys and brain imaging in monkeys and humans. Mirror neurons in the premotor cortex appear to be a part of the neuronal systems controlling motor actions. To advance further, it will be particularly important to find neural substrates of the psychological concepts of body schema and motor schema. Complex motor actions such as an actor’s performance on the stage typically integrate movements with perceptual and conceptual behavior. This indicates that neural mechanisms for motor actions must involve both motor and cognitive domains, as discussed in the next chapter.
In
Chapter 16
, “
Motor Actions and Tool Use
,” we examined the roles of the cerebellum in the control of complex motor actions in primates. We are now ready to extend our consideration to an even higher level of control; that for mental activity in humans. There is an increasing body of evidence (both experimental and theoretical) that supports the presumed unique roles of the cerebellum in our thought processes in which we manipulate ideas and concepts instead of moving body parts.
Here, we conceive a neural system for mental activities as an analogy of that for voluntary motor control and action control (
Chapters 13
–
16
). First, the prefrontal cortex is postulated to be the controller. The purpose is to orchestrate thought and action in accordance with (1) internal goals (
Miller and Cohen, 2001
), (2) the executive function required for the conscious control of thought and action (
Happaney et al., 2004
), and (3) the supervisory activating system for working memory (see following text). Second, controlled objects are represented in the temporoparietal cortex, which will be defined later in terms of mental models and schemata. Third, the neocerebellum provides cerebellar internal models (forward and inverse) of the controlled objects represented in the temporoparietal cortex. Thus, the prefrontal cortex, temporoparietal cortex, and neocerebellum conjointly constitute a control system for mental activities. In support of these views, the co-activation of these three structures corresponding to the controller, controlled object, and internal model, respectively, has been demonstrated in an increasing number of cases. For example, in a verb-to-noun conversion task, the prefrontal cortex and Wernicke’s area in the temporoparietal cortex (both of the left side) and the cerebellum (on the right side) have been shown to be co-activated (
Fiez et al., 1996
).
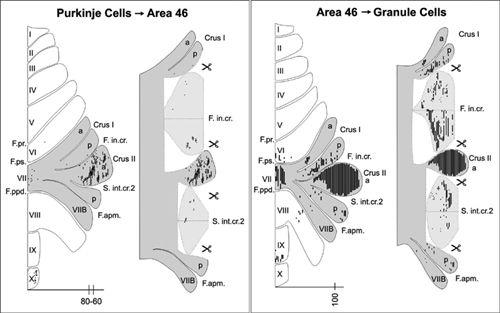
Kelly and Strick (
2003
) provided anatomical substrates for involvement of the cerebellum in cognitive function using viral tracing techniques in nonhuman primates (for review, see
Ramnani, 2006
;
Strick et al., 2009
). They revealed a loop connection between the prefrontal cortex (area 46) and the cerebellar hemispheres (crus I and crus II of the ansiform lobule) (
Figure 53
). In human subjects, Krienen and Buckner (
2009
) used functional connectivity MRI (fcMRI) to identify four topographically distinct frontocerebellar circuits that targeted the (1) motor cortex, (2) dorsolateral prefrontal cortex, (3) medial prefrontal cortex, and (4) anterior prefrontal cortex. A direct comparison between the right- versus left-side frontal regions revealed contralateral lateralization in the cerebellum for each of the segregated circuits. Overall, the extent of the cerebellum associated with the prefrontal cortex included a large portion of the posterior hemispheres associated with non-motor functions.
Figure 53. Input-output organization of the cerebellar loops with cortical area 46.
(Left) Distribution of Purkinje cells (small dots) that project to area 46. (Right) Distribution of granule cells (fine lines) that receive input from area 46. The shaded areas on the flattened surface maps (diagrams on the left in each panel) are unfolded on the right side of each panel to show the distribution of labeled neurons in the relevant cerebellar cortical fissures. Scale bars, 15 millimeters, (ignore the numbers on the X axes, which indicate the positions of two sections illustrated in another figure). (From
Kelly and Strick, 2003
.)
Anatomical connections have also been shown to exist between the temporoparietal cortex and the cerebellum. In nonhuman primates, the posterior parietal areas of the parietal cortex were shown to receive inputs from the ventral portions of the dentate nucleus in the cerebellar hemisphere (
Dum and Strick, 2003
) and the anterior intraparietal area of the posterior parietal cortex received inputs from a broad area of the dentate nucleus (
Clower et al., 2005
). In a human fcMRI study, connections were shown between the inferior temporal cortex and regions in the cerebellum (
Krienen and Buchner, 2009
). Because the inferior temporal cortex receives few anatomical connections from the pons, these latter connections must be mediated indirectly.
Sensory information about a stimulus is first represented in the primary sensory area of the cerebral cortex. Then, while being transferred through sensory association cortices, its mental representation is formed and eventually reaches the level of conscious awareness. For example, “binocular rivalry” occurs in the anterior part of the infratemporal cortex where neurons can behave in accordance with conscious experiences, but not with what is actually sensed by the retina (
Sheinberg and Logothetis, 1997
). Wilder G. Penfield (1891–1976) demonstrated that electrical stimulation of certain areas of the human cerebral cortex invokes various mental representations, even an episode that occurred in the past (
Penfield and Rasmussen, 1950
;
Penfield and Perot, 1963
).
A mental representation of complex integrated information may constitute a “mental model” that Craik (
1943
) defined as a psychological substrate of a mental representation of both real and imaginary situations. A mental model is a small-scale version of reality, which the mind constructs and uses to reason, explain, and anticipate a future event. In other words, mental models are representations of images, concepts, and ideas. No information is yet available, however, on just how mental models are expressed as specific activity in a cortical circuit. The concept of “schema” introduced by Jean Piaget (
1951
) is also close to that of mental models (recall
Chapter 1
, “
Neuronal Circuitry: The Key to Unlocking the Brain
”). His idea was that a child develops a schema that helps in interpreting and understanding the world. It includes both a category of knowledge and the process of obtaining that knowledge. An assembly of mental models constitutes an internal world that each individual’s conscious awareness can access.
It seems likely that a mental model is encoded in a neuronal circuit of the cerebral association cortex. Neurophysiological mapping in monkeys revealed activities related to the perception of various objects (e.g., dolls and artificial fruits) in the
temporoparietal association areas (
Tanaka, 1996
;
Tsunoda et al., 2001
). Neuroimaging revealed that neuronal activity associated with motor imagery required to simulate a movement involves activity occurring simultaneously in the premotor cortex, parietal association area, and cerebellum (
Jeannerod and Frank, 1999
). Such activity in the parietal cortex may represent a mental model of the movement. However, a more complex way of representing perception and behavior has been suggested (
Hesslow, 2002
). The brain may simulate motor behavior by activating motor structures, as during an overt action but suppressing its actual execution. Similarly, the brain may simulate perception by the internal activation of the cerebral sensory cortex, as during the normal perception of external stimuli. These considerations suggest that mental models elicit the perceptual simulation of the normal consequences of overt actions. Here, we consider that Craik’s mental model and Piaget’s schema are useful explanatory concepts, giving a psychological counterpart of the cerebral cortical models we assume to operate in the control system mechanisms for cognitive functions.
We assume that during repeated thoughts the cerebellum forms an internal model of the cerebral cortical mental model of these thoughts. A cerebellar forward and inverse model may copy essential properties of such a mental model. However, in contrast to a movement in the physical domain that can be computationally represented in an internal model, it remains unknown how a mental model having an abstract conceptual nature is represented in a neuronal circuit of the cerebral cortex and is copied by a circuit in the cerebellum. Mechanisms have been discussed for forming a computable mental model residing in the cerebral cortex (
Johnson-Raird, 1983
). Indeed, an episodic-like (event) memory or semantic-like (fact) memory is encoded in a neuronal circuit of the medial temporal lobe (see
Miyashita, 2004
). After it is encoded in a cortical network, it may be transferred by error learning to a network of the cerebellar cortex. To determine just how this is done is a great challenge in contemporary computational neuroscience.
Wolpert et al. (
2003
) explored the computational parallels between the processes that occur in motor control and in social interaction. They examined how models of motor control, such as the HMOSAIC (consisting of several layers of MOSAIC;
Haruno et al., 2003
), could be used for action observation, imitation, social interaction, and Theory of Mind (see
Section 7
). They suggested that models of motor control provide an efficient mechanism for performing the computations needed in social interaction.
In accordance with our daily life experiences, we assume that thought is performed through two parallel processes—one explicit and the other implicit. When we try to solve a novel problem that requires mentation, we initially devote strenuous conscious effort to the issue and continue to think about its solution. Later, we may recall the problem from time to time, but we are otherwise unaware that we are still trying to solve the problem. Yet, the thought of how this is to be done appears to proceed implicitly because the sought solution may be derived suddenly without obvious conscious effort to obtain it.
We further assume that the implicit process for the manipulation of our thoughts proceeds in the cerebellum, whereas the explicit process is undertaken in the cerebral cortex. Various forms of motor learning involving the cerebellum are indeed executed implicitly. Whereas electric stimulation of the cerebral cortex evokes conscious experiences in humans (
Penfield and Perot, 1963
), that of the cerebellar cortex causes no more than increased alertness and reduced depression and anxiety (
Riklan et al., 1976
). TMS of the cerebellum does not evoke conscious experience (
Koch et al., 2006
). These observations support the assumption that thought processes in the cerebellum do not reach the level of conscious awareness. We do not assume, however, that all events processed in the cerebral cortex reach conscious awareness because brain imaging has shown repeatedly that self-unrecognized activities take place in many cerebral cortical areas.