On Monsters: An Unnatural History of Our Worst Fears (30 page)
Read On Monsters: An Unnatural History of Our Worst Fears Online
Authors: Stephen T. Asma

Just as homologies demonstrate ancestry rather than divine ideas, so, too, some monsters demonstrate ancestry rather than potential new species. Recall Darwin’s description of humans born with werewolf syndrome, long hair covering their entire body and face. In
The Descent of Man
he considered cases of werewolf syndrome together with humans who are born with tails and suggested that both kinds of monsters are evidence that human beings are descended from animal ancestors. He included them in a battery of substantiating phenomena, all grouped together in chapter i, “The Evidence of the Descent of Man from Some Lower Form.” He offered the anomalous case of a tailed human as an atavism of our prehuman ancestral connection to primates and therefore a compelling bit of evidence for evolution. But the human werewolf monster is “a more curious case.” Darwin argued that the hairy human anomaly is connected to a completely normal state of intrauterine fetal development. Around sixteen weeks after conception, all humans start to form fine hairs covering the body, called
lanugo
(Latin for “downy”). Shortly before birth the fetus sheds this lanugo and swallows it; it mixes with bile and mucus and becomes part of the meconium (the first bowel movement of the baby). Darwin didn’t know some of these facts, but he did know that lanugo was part of normal embryological processes. And he suggested that perhaps human werewolf syndrome was a case of “arrested development,” a tera-tological retention of a normally abandoned hair growth process. Darwin did not tarry on this topic, leaving it (placed in the same chapter with other “evidence of descent”) as merely suggestive of evolution. But strictly speaking, if the werewolf syndrome is a monstrous arrested development of embryological process, then it is
not
really evidence of ancestral descent in the same way that human tails might be.
In the case of both hairy monsters and tailed monsters, Darwin recognized the importance of teratologies, but not as sources of new evolutionary pathways. They are not causes; they are either vestigial remnants that demonstrate the
fact
of evolution, or they are cases of arrested development that reveal more to us about the mysteries of embryology.
Despite Darwin’s reining in of monsters, they remained provocative for many biologists, and more than one theory of evolution vied for allegiance in the late nineteenth and early twentieth century.
16
The botanist Hugo de Vries (1848–1935), for example, proffered a popular alternative to Darwinism called the “mutation theory of evolution.” “It must be obvious,” de Vries wrote in
Species and Varieties: Their Origin by Mutation
in 1904, “that this theory of natural selection leaves the question as to how the changes themselves are brought about, quite undecided. There are two possibilities, and both have been propounded by Darwin. One is the accumulation of the slight deviations of fluctuating variability, the other consists of successive sports or leaps taking place in the same direction.”
17
De Vries, who developed an early genetics hypothesis, performed breeding experiments on plants and decided that Darwin had been too hasty to dismiss the role of macromutational jumps.
A monstrous plant, the Nepaul barley (a variety of
Hordeum vulgare
), came under de Vries’s careful study, and he drew significant conclusions from its morphology. In the botanical world, flowers rarely grow directly out of the leaf of a plant, but Nepaul barley not only had the odd case of a flowered leaf but also a second, smaller floret growing above the first. This freakish morphology had no use, according to de Vries, and constituted “perhaps the most obviously useless structure in the whole vegetable kingdom.” Nevertheless, he explained, the mutation was as completely hereditary as any of the most elegant adaptations in nature. “Therefore,” he concluded, “it is one of the most serious objections to the hypothesis of slow and gradual improvements on the sole ground of their usefulness.”
18
This idea, that evolution occurs by sudden leaps without intermediaries, found its strongest supporter in Richard Goldschmidt (1878–1958). A controversial biologist, Goldschmidt made his reputation first in Germany by working on genetics and sex differentiation in gypsy moth varieties, and then in the United States (after fleeing the Nazis) by working on fruit fly mutations. His objections to the emerging “Mendelian-chromosomal theory” of genetics caused him to miss the boat of the dominant neo-Darwinian synthesis of natural selection and Mendelian genetics, called the
new synthesis
. But his mutation experiments led him to emphasize the importance of development, timing, rates of reaction, and function in ways that make some aspects of his biology seem more prescient than his peers acknowledged.
19
Goldschmidt coined the phrase “hopeful monsters” to refer to macromutational jumps that might, in time, form successful new
species. The idea of viable large-scale saltation was completely out of step with the neo-Darwinian biology of the 1930s and 1940s.
20
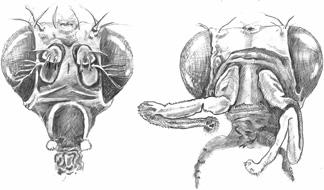
Compare the two heads of these fruit flies. On the left is a normal specimen; on the right the mutant fly has grown legs in the place of antennae. Pencil drawing by Stephen T. Asma © 2008.
Imagine a fruit fly (
Drosophilia melanogaster
) with arms growing in place of wings, or a fly whose wings were transformed into halteres (the balancing appendages on either side of the body), or a fly with legs growing in place of antennae (aristapedia). These and many other monstrosities increased in frequency when Goldschmidt shocked his embryonic flies with temperature changes. In fact, he found that mutations increased by six times if he administered such shocks. Now admittedly, a monster with legs in place of antennae is not terribly “hopeful,” but the basic jump of macromutation revealed by such experiments led him to suggest that hidden building mechanisms, such as segmentation, could somehow shift and thereby alter the organism’s phenotype. To use an analogy, if you think about micromutations as small-scale changes in the spelling of words on this page, Goldschmidt’s macromutations were more like transformations in the rules of grammar itself. Tweaking the grammar produces more dramatic effects in the overall result. Simple micromutations could not, in Goldschmidt’s view, rack up the major transformations that evolution seemed to require, whereas mutations in the grammar of development (eventually called homeotic mutations) seemed a better candidate.
When biologists want to understand an animal, they must
reverse-engineer
it. Like a mechanic who wants to learn how an unfamiliar engine came to be built, the biologist must infer the causal mechanisms that shaped the anatomy of an individual or a population. This can be exceedingly difficult to do, especially when very different causal stories are consistent with the animal’s present morphology. Is the particular shape of a fish’s lobe fin entirely the result of a strong environmental selection pressure, or is it best understood as the effect of internal laws of growth? By analogy, if one is reverse-engineering an unfamiliar vehicle, it may be the case that the oddly shaped exhaust pipe was developed as an adaptation for occasional submersion in water, but it might also be shaped this way simply because an enlarged catalytic converter forced the pipe into a unique position, and its successful submersion ability is just an accidental consequence.
Because the morphological traits of an animal are usually useful and functional in a specific environment, it is hard to determine the internal nonfunctional material laws that may have shaped the traits independently of environment pressure. The developmental biologist Pere Alberch (1954–1998) created a new movement in the 1990s when he proposed “that it may be advantageous to turn to non-functional, grossly maladapted, teratologies when studying the properties of internal factors in evolution.”
21
Because monstrous deviations from normal development are usually poorly adapted when compared with their parents, they will be selected against. “This is a useful property because if, in spite of very strong negative selection, teratologies are generated in a discrete and recurrent manner, this order has to be a reflection of the internal properties of the developmental system.” Like Goldschmidt, Alberch experimented with fruit flies. He found that the imaginal disks (tissues in the larval fly) could be manipulated to produce monsters, but the transformations were not chaotic: “For example, if a prospective genital cell makes a mistake, it will most likely produce a leg or a head-antenna tissue type…. It will never give rise to a wing or a thorax.” From these and other experiments Alberch concluded that mutations, whether natural or lab-induced, undergo a finite set of transformations. These limited trajectories, or developmental patterns, were not, he argued, sufficiently appreciated by neo-Darwinians, who tended to see mutation as unpredictable and almost infinitely potential.
In humans, for example, we have many cases of two-headed teratologies, but not three-headed. There is no good evidence of a single body axis that trifurcates at the anterior end into three heads.
22
Alberch surveyed
the historical reports and concluded that the only seemingly reputable report of a three-headed child (from the Italian physicians Reina and Galvagni) appeared to be a case of a conjoined twin with two distinct spinal chords and a further parasite head built on the axis of one of the doubled spines. Double monsters (conjoined twins) divide and branch in the same three or four discernable ways, whether they are men, cows, or fish. Alberch also pointed out that cyclops teratologies exhibit the same morphological aspects, regardless of the particular species or higher taxa in which they occur. Monsters come to the rescue for biologists who want to isolate internal causes, but Alberch saw himself as returning the favor, raising monsters from their demoted Darwinian status. He claimed that his interest in teratology “contrasts with the bad reputation that monsters have traditionally enjoyed in the Darwinian literature.” From a Darwinian perspective, teratologies are not adaptive variations, so they are “evolutionary dead ends” and “unworthy of study.” But Alberch and his subsequent school of typologists treated monsters as “model systems to study the patterning generated by developmental properties.” This approach to monsters continues to pick up steam and flourish today, but Alberch claimed that he was not a proponent of the “hopeful monsters” thesis of biologists like Goldschmidt. “I do not contend,” Alberch said, “that teratologies are variation with evolutionary potential. In fact, I assume them to be lethal in most cases.”
Stephen Jay Gould extrapolated on Alberch’s research and showed how teratology can be profitably linked to evolutionary questions.
23
Consider the pentadactyl hand of tetrapods. Gould pointed out that the earliest known tetrapods, from the late Devonian period (390–340 million years ago), were incomplete fossils, but paleontologists in the 1930s reconstructed their limbs as five-digit forms (assuming the orthodox view of pentadactyl homology). Surprisingly, however, more recent paleontological discoveries have shown that the earliest tetrapods had seven and sometimes even eight toes and fingers. It appears now that five digits was not an original vertebrate structure, but probably represents a later stabilization of form, one that still sits at a significant branching limb of the phylogenetic tree. But then Gould introduced some of Alberch’s teratology to interesting ends.
24
Before Alberch’s research it was thought that the formation of the forelimb followed a central axis, with digits forming outward from some theoretical middle-line origin. Alberch showed that the order of the limb and digit formation followed a pattern of construction from back to front: the pinky finger is built first and the thumb is last (in the feet, the little toe is first and the big toe is last).
25
This was surprising and counterintuitive. Gould pointed out that, under Alberch’s revision, “the array of digits
becomes a sequence of timing: Spatial position is a mark of temporal order. Back equals old; front is young.” Confirmation of this theory came from independent phenomena suggesting that many animals who grow extra digits (monstrous polydactyls) do so by failing to shut off the normal sequence, thereby adding an extra digit after the “youngest” digit.
26
