Hen’s Teeth and Horse’s Toes (2 page)
Read Hen’s Teeth and Horse’s Toes Online
Authors: Stephen Jay Gould

ALFRED, LORD TENNYSON
, never known for egalitarian perspectives, had this to say about the relative merit of the sexes:
Woman is the lesser man, and all
thy passions, matched with mine,
Are as moonlight unto sunlight, and
as water unto wine.
The couplet may not represent Tennyson’s considered view, since the protagonist of “Locksley Hall” had lost his love to another and speaks these words during a grand poetic fit of sour grapes. Still, the literal reading—that women are smaller than men—would be accepted by most of us as a general fact of nature, not as a sexist trap. And most of us would therefore be wrong.
Human males are, of course, generally larger than human females, and most familiar mammals follow the same pattern (but see essay 11). Yet females are larger than males in a majority of animal species—and probably a large majority at that. For starters, most animal species are insects and female insects usually exceed their males in size. Why are males generally smaller?
One amusing suggestion was proposed in all seriousness just 100 years ago (as I discovered in the “50 and 100 Years Ago” column in
Scientific American
for January, 1982). A certain M. G. Delaunay argued that human races might be ranked by the relative social position of females. Inferior races suffered under female supremacy, males dominated in superior races, while equality of sexes marked races of middle rank. As collateral support for his peculiar thesis, Delaunay argued that females are larger than males in “lower” animals and smaller in “higher” creatures. Thus, the greater number of species with larger females posed no threat to a general notion of male superiority. After all, many serve and few rule.
Delaunay’s argument is almost too precious to disturb with refutation, but it’s probably worth mentioning that the paradigm case of a “higher” group with larger males—the mammals—is shakier than most people think (see Katherine Ralls in the bibliography). Males are larger in a majority of mammalian species, of course, but Ralls found a surprising number of species with larger females, spread widely throughout the range of mammalian diversity. Twelve of 20 orders and 20 of 122 families contain species with larger females. In some important groups, larger females are the rule: rabbits and hares, a family of bats, three families of baleen whales, a major group of seals, and two tribes of antelopes. Ralls further reminds us that since blue whales are the largest animals that have ever lived, and since females surpass males in baleen whales, the largest individual animal of all time is undoubtedly a female. The biggest reliably measured whale was 93.5 feet long and a female.
The sporadic distribution of larger females within the taxonomic range of mammals illustrates the most important general conclusion we can reach about the relative size of sexes: the observed pattern does not suggest any general or overarching trend associating predominance of either sex with anatomical complexity, geological age, or supposed evolutionary stage. Rather, the relative size of sexes seems to reflect an evolved strategy for each particular circumstance—an affirmation of Darwin’s vision that evolution is primarily the story of adaptation to local environments. In this perspective, we must anticipate the usual pattern of larger females. Females, as producers of eggs, are usually more active than males in brooding their young. (Such male tenders as sea horses and various mouth-brooding fishes must receive eggs directly from a female or actively pick up eggs after a female discharges them.) Even in species that furnish no parental care, eggs must be provided with nutriment, while sperm is little more than naked DNA with a delivery system. Larger eggs require more room and a bigger body to produce them.
If females provide the essential nutriment for embryonic or larval growth, we might ask why males exist at all. Why bother with sex if one parent can supply the essential provisioning? The answer to this old dilemma seems to lie in the nature of Darwin’s world. If natural selection propels evolution by preserving favored variants from a spectrum randomly distributed about an average value, then an absence of variation derails the process—for natural selection makes nothing directly and can only choose among alternatives presented. If all offspring were the xeroxed copies of a single parent, they would present no genetic variation (except for rare new mutations) and selection could not operate effectively. Sex generates an enormous array of variation by mixing the genetic material of two creatures in each offspring. If only for this reason, we shall have males to kick around for some time.
But if the biological function of males does not extend beyond the contribution of some essentially naked DNA, why bother to put so much effort into making them? Why should they, in most cases, be almost as big as females, endowed with complex organs, and quite capable of an independent life? Why should industrious bees continue to make the large and largely useless male creatures appropriately known as drones?
These questions would be difficult to answer if evolution worked for the good of species or larger groups. But Darwin’s theory of natural selection holds that evolution is fundamentally a struggle among individual organisms to pass more of their genes into future generations. Since males are essential (as argued above), they become evolutionary agents in their own right; they are not designed for the benefit of their species. As independent agents, they join the struggle in their own ways—and these ways sometimes favor a larger size. In many groups, males fight (literally) for access to females, and heavyweights often have an edge. In more complex creatures, social life may emerge and become ever more elaborate. Such complexity may require the presence and active involvement of more than one parent in the rearing of offspring—and males gain a biological role transcending mere stud service.
But what of ecological situations that neither favor battle nor require parental care? After all, Tennyson’s most famous biological line—his description of life’s ecology as “Nature, red in tooth and claw”—does not apply in all, or most, cases. Darwin’s “struggle for existence” is a metaphor and need not imply active combat. The struggle for genetic representation in the next generation can be pursued in a variety of ways. One common strategy mimics the motto of rigged elections: vote early and vote often (but substitute “fornicate” for “vote”). Males who follow this tactic have no evolutionary rationale for large size and complexity beyond what they need to locate a female as quickly as possible and to stick around. In such cases, we might expect to find males in their minimal state, a status that might have become quite general if evolution worked for the good of species—a small device dedicated to the delivery of sperm. Nature, ever obliging, has provided us with some examples of what, but for the grace of natural selection, might have been my fate.
Consider a species so thinly spread over such a broad area that males will rarely meet at the site of a female. Suppose also that females, as adults, move very little if at all: they may be attached to the substrate (barnacles, for example); they may live parasitically, within another creature; or they may feed by waiting and luring rather than by pursuit. And suppose finally that the surrounding medium can easily move small creatures about—as in the sea, with its currents and high density (see M. Ghiselin’s book,
The Economy of Nature and the Evolution of Sex
, for a discussion of this phenomenon). Since males have little impetus for literal battle, since they must find a stationary female, and since the medium in which they live can provide (or substantially aid) their transport, why be large? Why not find a female fast when still quite small and young and then hang on as a simple source of sperm? Why work and feed, and grow large and complex? Why not exploit the feeding female? All her offspring will still be 50 percent you.
Indeed, this strategy is quite common, although little appreciated by sentient mammals of different status, among marine invertebrates that either live at great depth (where food is scarce and populations very thinly spread), or place themselves in widely dispersed spots that are hard to locate (as in many parasites). Here we often encounter that ultimate in the expression of nature’s more common tendency—females larger than males. The males become dwarfs, often less than one-tenth the length of females, and evolve a body suited primarily for finding females—a sperm delivery system of sorts.
A species of
Enteroxenos
, for example, a molluscan parasite that lives inside the gut of sea cucumbers (echinoderms related to sea urchins and starfishes), was originally described as a hermaphrodite, with both male and female organs. But J. Lutzen of the University of Copenhagen recently discovered that the male “organ” is actually the degenerated product of a separate dwarf male organism that found the parasitic female and attached permanently to her. The female
Enteroxenos
fastens herself to the sea cucumber’s esophagus by a small ciliated tube. The dwarf male finds the tube, enters the female’s body, attaches to it in a particular place, and then loses virtually all its organs except, of course, for the testes. After a male enters, the female breaks its tubular connection with the sea cucumber’s esophagus, thereby obliterating the pathway of entrance for any future males. (A strict Darwinian—I am not one—would predict that the male has evolved some device to break or cause the female to break this tubular connection, thereby excluding all subsequent males and assuring its own paternity for all the female’s offspring. But no evidence yet exists for or against this hypothesis.)
As long as such an uncomfortable phenomenon resides with unfamiliar and “lowly” invertebrates, male supremacists who seek pseudosupport from nature may not be greatly disturbed. But I am delighted to tell a similar story about one group of eminently suited vertebrates—deep-sea anglerfishes of the Ceratioidei (a large group with 11 families and nearly 100 species).
Ceratioid anglerfishes have all the prerequisites for evolving dwarf males as sperm delivery systems. They live at depth in the open ocean, mostly from 3,000 to 10,000 feet below the surface, where food is scarce and populations sparse. Females have detached the first dorsal fin ray and moved it forward over their capacious mouth. They dangle a lure at the tip of this spine and literally fish with it. They jiggle and wave the lure while floating, otherwise immobile, in the midst of the sea. The related shallower-water and bottom-dwelling anglerfishes often evolve elaborate mimetic structures for their lures—bits of tissue that resemble worms or even a decoy fish (see essay 3 in
The Panda’s Thumb
). Ceratioids live well below the depth that light can penetrate sea water. Their world is one of total ambient darkness, and they must therefore provide the light of attraction themselves. Their lures glow with a luminescence supplied by light glands—a death trap for prey and, perhaps, a beacon for dwarf males.
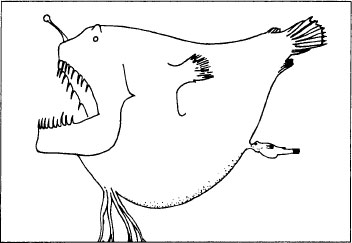
A male anglerfish (lower right), about one-and-a-half inches long, embeds itself into a ten-inch female of the same species.
REPRINTED FROM NATURAL HISTORY
.
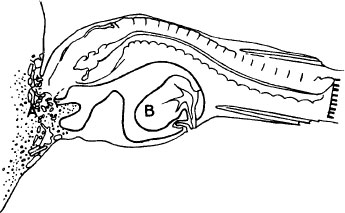
A simplified cross section shows a male anglerfish attached to a female. The two fish share tissue (A), and the male’s testis (B), has become enlarged.
REPRINTED FROM NATURAL HISTORY
.
In 1922, B. Saemundsson, an Icelandic fisheries biologist, dredged a female
Ceratias holbolli
, 26.16 inches in length. To his surprise, he found two small anglerfish, only 2.03 and 2.10 inches long, attached to the female’s skin. He assumed, naturally, that they were juveniles, but he was puzzled by their degenerate form: “At first sight,” he wrote, “I thought these young ones were pieces of skin torn off and loose.” Another oddity puzzled him even more: these small fish were so firmly attached that their lips had grown together about a wad of female tissue projecting well into their mouths and down their throats. Saemundsson could find no other language for his description but an obviously inappropriate mammalian analogy: “The lips are grown together and are attached to a soft papilla or ‘teat’ protruding, so far as I can see, from the belly of the mother.”