Hen’s Teeth and Horse’s Toes (12 page)
Read Hen’s Teeth and Horse’s Toes Online
Authors: Stephen Jay Gould

Evolution, Lysenko charged, is almost an illusion in Vavilov’s scheme. It represents a mere playing out of inherited potentials, not the development of anything new. It expresses the bourgeois penchant for stability by depicting apparent change as a superficial expression of underlying constancy. According to Vavilov’s law, Lysenko charged,
New forms result not from the development of old forms, but from a reshuffling, a recombination of already existing hereditary corpuscles…. All the existing species existed in the past, only in less diverse forms; but every form was richer in potentialities, in its collection of genes.
Madness often displays a perverse but cogent reason in its own terms; and we must admit that Lysenko did identify and exploit the true weaknesses in Vavilov’s argument. Vavilov did underplay the creative role of environment, and his chemical analogy did betray a belief in prefigured potentiality as the source of later, and in some sense illusory, change. But Lysenko, who was also both a charlatan and a cruel polemicist, was equally undialectical (despite his protestations to the contrary) in viewing plants as putty before a molding environment.
Vavilov died in the name of a phony Lamarckism. He became a legitimate martyr in the West, but his ideas did not flourish as a result. The law of homologous series, the organizing theme of his evolutionary work, was ignored in the name of an overly strict Darwinism. Vavilov’s law did not directly contradict Darwinian principles, but its emphasis on constraints of inheritance and channeled variation fit poorly with the favored Darwinian theme of random variation and the guidance of evolutionary change by natural selection. It was therefore neglected and relegated to the shelf of antiquated theories that had implicated variation itself as a directing force in evolution. I have consulted all the founding documents of the “modern synthesis,” the movement that established our present version of Darwinism between the late 1930s and the 1950s. Only two mention Vavilov’s law of homologous series, each in less than one paragraph.
Yet I feel that in his imperfect way Vavilov had glimpsed something important. In more modern terms, new species do not inherit an adult form from their ancestors. They receive a complex genetic system and a set of developmental pathways for translating genetic products through embryology and later growth into adult organisms. These pathways do constrain the expression of genetic variation; they do channel it along certain lines. Natural selection may choose any spot along the line, but it may not be able to move a species off the line—for selection can only act upon the variation presented to it. In this sense, constraints of variation may direct the paths of evolutionary change as much as selection acting in its Darwinian role as a creative force.
I have found Vavilov’s views very helpful in reorienting my own thinking in directions I regard as more fruitful than my previous unquestioned conviction that selection manufactures almost every evolutionary change. In studying the relationship of brain size to body size, biologists find that brains increase only one-fifth to two-fifths as fast as bodies in comparisons of closely related mammals differing only (or primarily) in body size—adults within a single species, breeds of domestic dogs, chimpanzees versus gorillas, for example. For ninety years, the large literature has centered on speculations about the adaptive reasons for this relationship, based upon the (usually unstated) assumption that it must arise as the direct product of natural selection.
But my colleague Russell Lande recently called my attention to several experiments on mice selected over several generations for larger body size alone. As these mice increased in size across generations, their brains enlarged at the characteristic rate—a bit more than one-fifth as fast as body size. Since we know that these experiments included no selection upon brain size, the one-fifth rate must be a side product of selected increase in body size alone. Since the one-fifth to two-fifths rate appears again and again in diverse lineages of mammals, and since it may record a nonadaptive response of brains to selection for larger bodies within mammalian developmental systems, the parallel sets of races and species arrayed along the one-fifth to two-fifths slope in carnivores, rodents, ungulates, and primates are non-Darwinian homologous series in Vavilov’s sense.
In personal research on the West Indian land snail
Cerion
, my colleague David Woodruff and I find the same two morphologies again and again in all the northern islands of the Bahamas. Ribby, white or solid-colored, thick, and roughly rectangular shells inhabit rocky coasts at the edges of banks where islands drop abruptly into deep seas. Smooth, mottled, thinner, and barrel-shaped shells inhabit calmer and lower coasts at the interior edges of banks, where islands cede to miles of shallow water. The easiest, and usual, conclusion would view ribby shells on all islands as closely related and smooth shells as members of another coherent group. But we believe that the complex set of characters forming the ribby and smooth morphologies arise independently, again and again. On the islands of Little Bahama Bank, both ribby and smooth animals share a distinctive genital anatomy. On the islands of Great Bahama Bank, both ribby and smooth develop an equally distinctive, but different, kind of penis. The ecology of rocky versus calm coasts may select for ribby and smooth morphologies as adaptations, but the coordinated appearance of the half dozen distinctive traits of each morphology may represent a channeling of available variation to produce homologous series (ribby and smooth varieties) in different lineages (defined by genital anatomy).
A complete theory of evolution must acknowledge a balance between “external” forces of environment imposing selection for local adaptation and “internal” forces representing constraints of inheritance and development. Vavilov placed too much emphasis on internal constraints and downgraded the power of selection. But Western Darwinians have erred equally in practically ignoring (while acknowledging in theory) the limits placed upon selection by structure and development—what Vavilov and the older biologists would have called “laws of form.” We need, in short, a real dialectic between the external and internal factors of evolution.
Vavilov’s personal tragedy cannot be undone. But he has been rehabilitated in Russia, where the All-Union Society of Geneticists and Selectionists now bears his name. We who view him as a martyr and champion his name while ignoring his ideas would do well to reconsider the older non-Darwinian tradition that he represented. Combined with our legitimate conviction about the power of selection, the principle of homologous series (and other “laws of form”) might foster an evolutionary theory truly synthetic in its integration of development and organic form with a body of principles now dominated by ecology and the effects of selection upon single genes and traits.
I FREELY ADMIT
that the spotted, or laughing, hyena is not the loveliest animal to behold. Still, it scarcely deserved the poor reputation imposed upon it by our illustrious forebears. Three myths about hyenas helped to inspire the loathing commentary of ancient texts.
Hyenas, first of all, were regarded as scavengers and consumers of carrion. In his
Natural History
, Pliny the Elder (
A.D.
23–79) spoke of them as the only animals that dig up graves in search of corpses (
ab uno animali sepulchra erui inquisitione corporum
). Conrad Gesner, the great sixteenth-century cataloger of natural history, reported that they gorge themselves so gluttonously after finding a corpse that their bellies swell to become taut as a drum. They then seek a narrow place between two trees or stones, force themselves through it, and extrude the remains of their meal simultaneously at both ends.
Hans Kruuk, who spent years studying spotted hyenas on their home turf (the plains of East Africa), has labored to dispel these ancient myths (see his book
The Spotted Hyena
, University of Chicago Press, 1972). He reports that hyenas will scavenge when they get the opportunity. (Almost all carnivores, including the noble lion, will happily feast upon the dead product of another animal’s labor.) But spotted hyenas live in hunting clans of up to eighty animals. Each clan controls a territory and kills most of its own food—mainly zebra and wildebeest—in communal, nocturnal pursuit.
As a second insult, hyenas were widely regarded as hybrids. Sir Walter Raleigh excluded them from Noah’s ark since he believed that God had only saved thoroughbreds. Hyenas were reconstituted after the flood through the unnatural union of a dog and cat. In fact, the three living species of hyena form a family of their own within the order Carnivora. They are most closely related to the mustelids (weasels and their allies).
As a final, phony blot on their escutcheon, and in the unkindest cut of all, many ancient writers charged that hyenas were hermaphrodites, bearing both male and female organs. Medieval bestiaries, always trying to draw a moral lesson from the depravity of beasts, focused on this supposed sexual ambivalence. A twelfth-century document, translated by T. H. White, declared:
Since they are neither male nor female, they are neither faithful nor pagan, but are obviously the people concerning whom Solomon said: “A man of double mind is inconstant in all his ways.” About whom also the Lord said: “Thou canst not serve God and Mammon.”
But hyenas also had some formidable defenders against this particular calumny. Aristotle himself had declared in the
Historia animalium:
“The statement is made that the hyena has both male and female sexual organs; but this is untrue.”
Aristotle—and not for the first time—was right of course. But the legend had arisen for a good reason. Female hyenas are virtually indistinguishable from males. Their clitoris is enlarged and extended to form an organ of the same size, shape, and position as the male penis. It can also be erected. Their labia have folded up and fused to form a false scrotum that is not discernibly different in external form or location from the true scrotum of males. It even contains fatty tissue forming two swellings easily mistaken for testicles. Authors of the most recent paper on spotted hyenas found the appearance of males and females “so close that sex could only be determined with certainty by palpation of the scrotum. Testes could be located in the scrotum of the male compared with soft adipose tissue in the false scrotum of the female.”
British zoologist L. Harrison Matthews wrote the most extensive anatomical description of the hyena’s sexual anatomy in 1939. He described the peniform clitoris, emphasizing that it is no smaller than the male penis, is equally constricted to a single slitlike opening at the tip, and is as subject to erection as its male counterpart. He concluded his dry and precise pages of description with as forceful a statement of wonder as measured British scientific prose would allow: “It is probably one of the most unusual of the forms which the external orifice of the urogenital canal takes amongst female mammals.”
Harrison Matthews also investigated the interesting question of how hyenas do it, given a female orifice no larger than the slit of a male’s penis. “In the pre-pubertal state,” he writes, “these functions are obviously impossible, owing to the minute size of the opening.” But as the female matures the slit gradually lengthens and “creeps down round the ventral surface…travelling down the midline” until it forms an orifice 1.5 cm long and extending from the tip of the clitoris to its base. This lengthening of the slit and a subsequent enlargement of the nipples following pregnancy and parturition help distinguish older females from males. We can now understand the basis for ancient myths that hyenas were either simultaneous hermaphrodites (bearing male and female organs at the same time) or male for part of their life and then female.
Nature’s oddities cry out for explanation, and we therefore ask what advantages females gain from looking like males. Immediately, we come upon the other most striking oddity of hyena biology: females not only resemble males, they are also larger than males, contrary to the usual pattern in mammals, including humans (but see essay 1 for a discussion of the reverse pattern in most other animals). Females in Kruuk’s East African clans averaged 120 pounds in body weight versus 107 pounds for males. Moreover, they lead the clans in hunting and defense of territory and are generally dominant over males in individual contacts. Dominance is not merely a result of larger size because females also rank higher than larger males if the discrepancy in size is not too great.
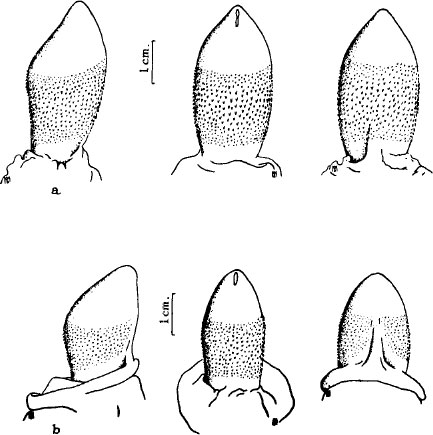
Similarity of male and female genitalia in the spotted hyena. Top row, views of the male penis. Bottom row, similar views of the female clitoris.
FROM HARRISON MATTHEWS
, 1939.
Although the female hyena’s assumption of what are usually male roles in mammals is probably related to its evolution of sexual structures that mimic male organs, the link between these phenomena is not immediately clear. It cannot have much to do with sexual performance itself for, if anything, the female “penis” is a hindrance to copulation until its opening enlarges and its form departs from that of the male.
Kruuk suggests that the strong mimicry arose in connection with a common behavior in hyenas called the “meeting ceremony.” Hyenas live in clans that defend territories and engage in communal hunting. But individuals also spend much of their time as solitary wanderers searching the landscape for carrion. To maintain cohesion in clans and to keep strangers away, hyenas must develop a mechanism for recognizing each other and reintegrating solitary wanderers into their proper clan.
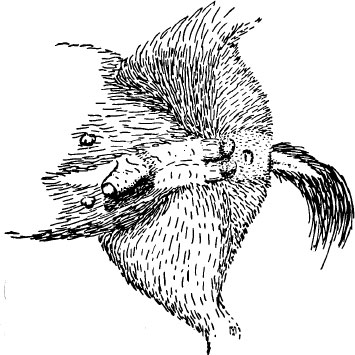
External genitalia of female spotted hyena, showing peniform clitoris and false scrotum.
FROM HARRISON MATTHEWS
, 1939.
When two hyenas of the same clan meet, they stand side to side, facing in opposite directions. Each lifts its inside hind leg, subordinate individual first, exposing either an erect penis or clitoris, one of the most vulnerable parts of the body, to its partner’s teeth. They then sniff and lick each other’s genitals for ten to fifteen seconds, primarily at the base of the penis or clitoris and in front of the scrotum or false scrotum.
Kruuk believes that the female clitoris and false scrotum evolved to provide a conspicuous structure serving for recognition in the meeting ceremony. He writes:
It is impossible to think of any other purpose for this special female feature than for use in the meeting ceremony…. It may also be, then, that an individual with a familiar but relatively complex and conspicuous structure sniffed at during the meeting has an advantage over others; the structure would often facilitate this reestablishment of social bonds by keeping partners together over a longer meeting period. This could be the selective advantage that has caused the evolution of the females’ and cubs’ genital structure.
Speculation about adaptive significance is a favorite, and surely entertaining, ploy among evolutionary biologists. But the question, “What is it for?” often diverts attention from the more mundane but often more enlightening issue, “How is it built?” In this case, speculations about adaptive significance have been in the literature for a long time, yet no one bothered to tread the obvious path for hypotheses of anatomical construction until 1979: What sexual hormones are maintained at what levels by female hyenas from conception to maturity? (See Racey and Skinner, 1979, in bibliography).
Racey and Skinner found, in short, that two androgens (male-producing hormones) had higher concentrations in testicles than in ovaries of adult spotted hyenas (scarcely surprising). Yet, when they investigated levels of the same hormones in blood plasma, they detected
no differences
between males and females. One female contained twin female fetuses, and both had about the same level of testosterone as adult females. Racey and Skinner therefore conclude “that high foetal androgen levels are responsible for the appearance of the male sexual facies in adult female spotted hyenas.”
Racey and Skinner affirmed their hypothesis by studying brown and striped hyenas, the other two species of the family Hyaenidae. Neither brown nor striped hyenas develop peniform clitorises or false scrotums. In both species, androgen levels in blood plasma are much lower for females than for males. (Aristotle, by the way, defended hyenas against the charge of hermaphroditism by correctly describing the genitalia of these other species—something of a dodge with respect to the spotted hyena, the source of the legend; but “the master of them that know” was right in any case.)
But why should high levels of androgenic hormones lead to the construction of false penises and scrotums? The animals that form them are still, after all, genetically female. How can female genes produce mimics of male structures, even in a milieu of high androgenic hormones? A look at the developmental basis of sexual anatomy resolves this dilemma.
Mammals share a common pattern for the embryology of sexual organs, and we may therefore use humans as an example. The early embryo is sexually indifferent and contains all precursors and structures necessary for the development of either male or female organs. After about the eighth week following conception, the gonads begin to differentiate as either ovaries or testes. The developing testes secrete androgens, which induce the development of male genitalia. If androgens are absent, or present at low levels, female genitalia are formed.
The internal and external genitalia develop in different ways. For internal genitalia, the early embryo contains precursors of both sexes: the Müllerian ducts (which form the Fallopian tubes and ovaries of females) and the Wolffian ducts (which form the vas deferens—the ducts that carry sperm from the testes to the penis—in males). In females, the Wolffian ducts degenerate and the Müllerian ducts differentiate; males develop by the opposite route.
The external genitalia follow a markedly different pattern. Individuals do not begin with two distinct sets of precursors and then lose one while strengthening the other. Rather, the different organs of male and female develop along diverging routes from the
same
precursor. The male’s penis is the same organ as the female’s clitoris—they form from the same tissues, are indistinguishable in the early embryo, and follow different pathways later. The male’s scrotum is the same organ as the female’s labia majora. The two lips simply grow longer, fold over and fuse along the midline, forming the scrotal sac.
The female course of development is, in a sense, biologically intrinsic to all mammals. It is the pattern that unfolds in the absence of any hormonal influence. The male route is a modification induced by secretion of androgens from the developing testes.
The mystery of male mimicry in female hyenas may be solved, in large part, by recognizing these fundamental facts of developmental anatomy. We know from the work of Racey and Skinner that female hyenas maintain high levels of androgenic hormones. We may therefore conclude that the striking and complex peculiarities of sexual anatomy in female spotted hyenas are simply, indeed almost automatically, produced by a single, underlying effect: the secretion of unusually large amounts of androgens by females.