Brilliant Blunders: From Darwin to Einstein - Colossal Mistakes by Great Scientists That Changed Our Understanding of Life and the Universe (22 page)
Authors: Mario Livio

In my conversation with Jack Dunitz, he recalled that Pauling had once told him something that summarized beautifully his attitude toward scientific research:
Jack, if you think you have a good idea, publish it! Don’t be afraid to make a mistake. Mistakes do no harm in science because there are lots of smart people out there who will immediately spot a mistake and correct it. You can only make a fool of yourself and that does no harm, except to your pride. If it happens to be a good idea, however, and you don’t publish it, science may suffer a loss.
Dunitz added that indeed the three-stranded structure did no harm, except to Pauling’s reputation. He commented further that Pauling had made enough major contributions that we should simply forgive and forget. I must say that I fully agree with the “forgive” part, but I actually think that we should
not
forget. As I have attempted to show, there are many insights that can be gained from analyzing such blunders by brilliant individuals.
Seeing Double
The rest of the story of the discovery of the structure of DNA has been told and retold numerous times, but the recently discovered correspondence of Francis Crick does shed some new light on the frantic activity that preceded the publication of the Watson and Crick model.
Pauling’s blunder served as the catalyst that convinced Bragg to allow Watson and Crick to go back to DNA modeling. Within a couple of weeks, Watson went to London, where Wilkins, also pleased with Pauling’s glitch, took the liberty of showing him Franklin’s famous photograph 51 of the B form of DNA (figure 14), without Franklin’s knowledge. Much ink has been devoted to the question of the ethical nature of this particular act. In my humble opinion, three main parts to this story deserve attention. First, there was apparently no problem with Wilkins himself having a copy of the photo (given to him by Gosling), since Franklin was about to leave King’s to work at Birkbeck College, and she had been informed by the lab director, Sir John Randall, that the results of all the DNA work belonged exclusively to King’s. Second, there is little doubt (in my mind at least) that Franklin should have been consulted before her
unpublished
results were shared with members of another laboratory. Finally, there is disagreement on whether or not Watson and Crick acknowledged Franklin’s contribution adequately in their paper. You can be the judge of that. They wrote,
“We have also been stimulated by a knowledge of the general nature of the unpublished experimental results and ideas of Dr. M. H. F. Wilkins, Dr.
R. E. Franklin and their co-workers at King’s College, London.” Be that as it may, the effect the photo had on Watson was dramatic:
The dark cross was the unmistakable sign of a helical structure. No wonder that, as he later described,
his “mouth fell open,” and his “pulse began to race.”
Watson and Crick spent the following weeks trying frantically to build models in which the bases would form the rungs of the helical ladder they had in mind. The first attempts were unsuccessful. Ignoring the clue from Chargaff’s ratios, Watson mistakenly thought that he should pair every base with its twin, forming rungs composed of adenine-adenine (A-A), cytosine-cytosine (C-C), guanine-guanine (G-G), and thymine-thymine (T-T). However, since the bases C and T were different in length from G and A, this created rungs of unequal lengths, which was inconsistent with the symmetric pattern exhibited in photograph 51. There was also the question of the bond between the two bases in each rung and between the rung and the “legs” of the ladder (which were supposed to be composed of sugars and phosphates). Here again Watson and Crick were heading the wrong way, but their office mate
Jerry Donohue came to the rescue. As a former student of Pauling’s, Donohue knew everything there was to know about hydrogen bonds. Donohue pointed out to Watson and Crick that even many textbooks had the hydrogen atoms in the wrong positions in thymine and guanine. Placing these atoms in their correct locations opened new possibilities for bonding the bases to each other. By shifting the bases in and out of other pairing possibilities (than the like-with-like), Watson suddenly realized that an A-T pair held together by two hydrogen bonds was identical to a G-C pair held similarly. The rungs became of equal length. Moreover, this pairing provided a natural explanation to Chargaff’s rules. Clearly, if A always paired with T, and G with C, then the numbers of A and T molecules in any section of DNA would be equal, and similarly for G and C. Another source of valuable information became available around that time via Max Perutz: a copy of Franklin’s report, written for a visit of the Medical Research Council biophysics committee to King’s.
From the symmetry of the crystalline DNA
described in that report, Crick concluded that the two strands of DNA were antiparallel—they ran in opposite directions.
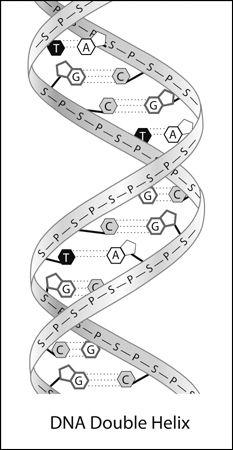
Figure 15
The resulting structure was the celebrated double helix, in which the two helical strands (the backbones) were made of alternating phosphates and sugars, with the paired bases attached to the sugars and making the rungs (
figure 15
). At this point, Watson and Crick were so convinced of the correctness of their model that they were eager to submit a short paper to
Nature
to announce it. Even before that, according to Watson’s by-now-famous description, Crick interrupted patrons’ lunchtime at the Eagle to make public that he and Watson had “discovered the secret of life.”
Figure 16
shows the spot in the Eagle where Crick made the announcement. On March 17, 1953, Crick sent a copy of the paper to Wilkins. One of the recovered documents in Crick’s “lost” correspondence is a draft of the letter that was to accompany the manuscript. Part of it reads:

Figure 16
Dear Maurice,
I enclose a draft of our letter. As it has not yet been seen by Bragg I would be grateful if you did not show it to anyone else. The object of sending it to you at this stage is to obtain your approval of two points:
a) the reference number 8 to your unpublished work.
b) the acknowledgement.
If you would like either of these rewritten, please let us know. If we don’t hear from you within a day or so we shall assume that you have no objection to their present form.
This draft and another one addressed to one of the editors of
Nature
(which apparently was never mailed) show that Crick and Watson
were at first under the impression that theirs was the only manuscript to be submitted at that time. Actually, the two groups at King’s submitted papers to
Nature
as well. In a brief note to Crick probably written on the same day, Wilkins says, “Herewith almost uncorrected draft. How should we refer to your note?” This accompanied a draft of Wilkins’s own manuscript. The third paper was by Rosalind Franklin and Raymond Gosling.
Once he realized the situation, Crick expressed his view that everyone should see everyone else’s manuscript: “It is not reasonable for letters to be sent jointly to
Nature
without having been read by all concerned. We want to see hers [Franklin’s], and I’ve no doubt she wishes to see ours.” Wilkins agreed. In a newly found letter dated “Mon.,” probably referring to Monday, March 23, he said, “We will post a copy of Rosy’s thing to you tomorrow,” adding, “Raymond and Rosy have your thing so everybody will have seen everybody else’s.”
Perhaps the most fascinating part of the new correspondence, however, is related to Pauling. First, Crick expressed his displeasure with the fact that Franklin might want to see Pauling on his forthcoming visit to England. “It is not impossible,” he wrote to Wilkins, “that she might consider turning over the experimental data to Pauling. This would inevitably mean that Pauling would prove the structure and not you.” To which Wilkins responded with irritation:
“If Rosy wants to see Pauling, what the hell can we do about it? If we suggested it would be nicer if she didn’t that would only encourage her to do so. Why is everybody so terribly interested in seeing Pauling . . . Now Raymond [Gosling] wants to see Pauling too! To hell with it all.” This exchange is a perfect demonstration of the awe that Pauling continued to inspire even at one of the lowest moments in his career.
The April 25, 1953, issue of
Nature
contained three papers on the structure of DNA. First, there was
the landmark paper by Watson and Crick describing the double helix structure. The paper was just a little more than one page long, but what a page that was.
Watson and Crick started by acknowledging, “A structure for nucleic acid has already been proposed by Pauling and Corey. They kindly made their manuscript available to us in advance of publication.” However, they immediately added, “In our opinion, this structure is unsatisfactory.” They then concisely explained their “radically different structure,” consisting of “two helical chains each coiled around the same axis,” and, in particular, the “novel feature” of the structure, which is “the manner in which the two chains are held together by the purine and pyrimidine bases.”
Watson and Crick’s model immediately suggested a solution both to how the coding of genetic information is achieved and to the puzzle of how the molecule manages to copy itself.
The details were presented in a second paper, published just five weeks after the first, in which Watson and Crick proposed the mechanism underlying the genetic code: “The phosphate-sugar backbone of our model is completely regular, but any sequence of the pairs of bases can fit into the structure. It follows that in a long molecule many different permutations are possible, and it therefore seems likely that
the precise sequence of the bases is the code which carries the genetical information
[emphasis added].” The message was clear: The coding of the genetic instructions that are needed to create, say, an amino acid, is contained in the specific sequence of bases in the rungs. For instance, the sequence C-G followed by G-C and then by T-A codes for forming the amino acid arginine, while G-C followed by C-G and then by T-A codes for alanine. The copying is done (precisely as anticipated abstractly by Pauling in 1948) by “unzipping” the double helix ladder at its center, producing two halves, each containing a leg and one-half of each one of the rungs. Because the sequence of bases in one chain automatically determines the sequence of the bases in the other (since the partner of T is always A, and that of G is always C), it is clear that one-half of the molecule contains all the information needed for constructing the whole molecule. For instance, if the sequence of bases along one chain of DNA is TAGCA, then the complementary sequence in the other
chain must be ATCGT. This way, two new complete ladders can be generated from the original one and, hence, copying of the DNA molecule is accomplished.
In their first paper, Watson and Crick did not spell out the copying mechanism, but they remarked laconically, “It has not escaped our notice that the specific pairing we have postulated immediately suggests a possible copy mechanism for the genetic material.”
Crick explained later that this enigmatically economical sentence (which has been labeled “coy” by some historians of science) was, in fact, a compromise between his own desire to discuss the genetic implications in the first paper and Watson’s concern that the structure might still be wrong. The statement was, therefore, a simple claim to priority. The fact that Watson did still harbor doubts about the model is well documented in his contemporary letters.
As I have noted, two other papers in
Nature
accompanied the first paper by Watson and Crick.
One was by Wilkins, Alexander Stokes, and Herbert Wilson, in which they analyzed some of the X-ray crystallographic data and also presented evidence that the helical structure exists not just in isolated fibers but also in intact biological systems. In the years that followed, Wilkins and his colleagues, and also Matthew Meselson, Arthur Kornberg, and others, did much work to confirm in detail the Watson and Crick model and their conclusions.