The Cannabis Breeder's Bible (31 page)
Read The Cannabis Breeder's Bible Online
Authors: Greg Green

Inbreeding increases homozygosity and crossbreeding maximizes heterozygosity. Crossbreeding takes advantage of hybrid vigor, which results in bursts of fertility and growth. The differences between the two parents are what result in hybrid vigor. In a large population where unrelated breeds of a species mate, the offspring in the first generation will be intermediate in characteristics to the parents and some will express the dominant phenotypes of both strains. Population means are deduced from the differences in gene frequencies among populations. We will have a look at these intermediate characteristics and how they can combine to produce hybrid vigor in a moment.
HYBRID VIGOR
Heterosis is a function of the square of the difference in gene frequency multiplied by the dominance deviation. This hypothesis states that heterosis is caused by a superior manifestation of “deleterious genes” when homozygosity is amplified and the heterozygote terminated somewhere in between the two corresponding homozygotes. It can also be supposed that heterosis occurs because of the termination of deleterious recessives contributed by one of the parents in the heterozygous F1 cross. All of this will make more sense in a moment when we see the possible combinations of genes in a population with our hybrid vigor model.
So what is a deleterious recessive condition? Deleterious alleles reduce the likelihood of an individual’s survival while safe alleles (non-deleterious) increase the chances of an individual’s survival (basically bad traits versus good traits that have an impact on the plant’s survival). If the recessive trait is a weak trait then fewer offspring will be produced to contribute to the population. Plants with deleterious alleles are usually eliminated quicker than healthier individuals.
Deleterious alleles exist and increase in a non-uniform population because of non-random mating factors. A deleterious recessive trait will generally cause a plant to use more resources in an effort to balance out the problem. Sometimes a deleterious recessive trait may cause a plant to grow taller with long internodes, naturally causing it to receive more light and it may dominate the smaller plants which have less non-deleterious recessive alleles. So in effect the deleterious gene has dominated the non-deleterious gene by default of its other dominant values. However this is rare and in most cases the number of deleterious alleles is kept low by a process of natural selection.
Breeders have been able to sustain plants that should not have survived in the wild because of their deleterious genes. This depends on what we consider a deleterious allele. Bad resistance to a certain type of pest can be treated as deleterious, yet indoors an IBL line may not encounter this pest and so the deleterious trait goes unnoticed. If this plant population were to exist in a locale where the pest is also present then the deleterious trait would be a disadvantage to the population and it might not continue. The deleterious allele will usually burden the plant in a population and it will use more energy and resources to survive. It uses more resources than plants with the non-deleterious genes for that trait and so is considered a waste of sorts in comparison. But plants with a deleterious allele may prove successful in dealing with a problem that a strain encounters. Again, variations are more adaptable. Recessive alleles are not classed as harmful or beneficial. The same goes for dominant alleles.
If we can understand these points and remember what we have learned about the recombination of cannabis genetic material then we should be able to understand hybrid vigor in the model below.
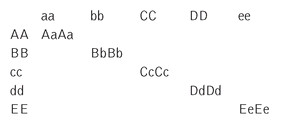
The most important element here is the consideration of a plant that is AaBbCcDdEe. Notice that none of the parents have this exact combination. The parents are AABBccddEE and aabbCCDDee. The first parent, AABBccddE E, has two recessives (cc, dd). The second parent, aabbCCDDee, has three recessives (aa,bb,ee). The offspring that has Aa,Bb,Cc,Dd,Ee is dominant for A,B,C,D and E. These A B C D E alleles are all present in this offspring and their dominance will be displayed in their respective phenotypes, which gives the plant its impression of hybrid vigor. This offspring is unique! Please remember, though, that we are using this model in the context of recessive and partially recessive deleterious alleles in respect to hybrid vigor.
10
You can see how a superior plant can emerge in a population via natural selection in hybrids. This is why it is important to grow large populations for selections when you are looking for that special mother plant that does better than all the rest. Usually the special mother can be attributed to hybrid vigor.
There is a lot of controversy about hybrid vigor because there is a back element to it that says that maybe the dominant allele is not the best allele for a new breeding environment. By breeding nonselectively in your environment you should help influence traits that suit that environment, with the subsequent removal of the deleterious alleles that do not suit your environment. The adaptable vigor of the F1 hybrid cross will decrease as we inbreed more.
“Over-dominance” is a term also used to explain hybrid vigor. True over-dominance occurs when there are multiple alleles for the same trait brought to the table in numbers of more than two. If nonrandom mating occurs then exceptional multiple hybrid variations for the same trait may be produced. As you can guess the more hybrids that are created the more recombinations will occur and the greater the chances of producing a vigorous plant that is seeking resolution in its environment.
In short, breeders develop new and better cannabis genetics by reshuffling the genetic code rather than inbreeding. Inbreeding stabilizes a strain by locking down homozygous traits.
AN INTRODUCTION TO THE SEXUAL EVOLUTION OF CANNABIS
Selfed plants usually produce more seeds because of selective pressures. Having pollen readily available is part of reproductive assurance.
In 1932 John Burton Haldane published “The Causes of Evolution.” Haldane made the discovery that there was a higher probability of encouraging a fixation of beneficial alleles for adaptability in selfing rather than outcrosses because there is a greater forwarding of these alleles to the overall selection process. This process, known as Haldane’s Sieve, also has negative effects because it forfeits long-term adaptability in favor of short-term advantages. This argument is known as “adaptedness,” from the adaptability hypothesis founded by S. K. Jain in 1976 in his publication “The Evolution of Inbreeding in Plants.”
In 1957 Dr. G. Ledyard Stebbins published “Regularities of Transformation in the Flower,” “Self-Fertilization and Population Variability in the Higher Plants,” and “The Inviability, Weakness and Sterility of Inter-Specific Hybrids” among many other important botany papers that year. Stebbins suggests that the evolution of selfing is an amalgamation of preliminary reproductive assurances followed by adaptability. Stebbins discovered that it is beneficial for a species to preserve welladapted genotypes over long periods of time if the environment suits the population.
We have seen both advantages and disadvantages to selfing. In short, selfing increases reproductive assurance but can decrease long-term success (As a note, we are no longer considering the fact that sinsemilla crops from non-selfed populations are what the cannabis grower is looking for. Here, rather, we are considering the evolutionary processes of the dioecious and monoecious conditions in cannabis). In their 1985 publication “The Evolution of Self-Fertilization and Inbreeding Depression in Plants,” R. Lande and D. W. Schemske found that there is actually a disorderly selection for selfing rates in wild plant populations which prevents the hermaphrodite condition from being more widespread. Both selfing (autogamy) and normal reproduction (allogamy) are styles of mating practices and are dependent on the fitness of the plant. Self-fertilization in mostly outcrossed populations will cause high levels of inbreeding depression because of higher genotype frequencies of recessive homozygotes which, in the absence of strong inbreeding, will not be purged. Thus, inbreeding in mostly outcrossed populations will tend to promote the maintenance of outcrossed systems and not selfing.
In 1996 M. Schierup and F.B. Christiansen in their publication “Inbreeding and Outbreeding in Plants” and in 1997 M. Fischer and D. Matthies in their publication “Mating Structure, and Inbreeding and Outbreeding Depression in the Rare Plant Gentianella Germanica (Gentianaceae)” considered that in contrast if we look at mostly selfing populations, the plants are highly adapted to their particular forte. Outcrossing, either with plants from another population by migration or from within the same population, can cause “Outbreeding Depression” where the offspring of the outcrosses are less fit than the offspring of selfing. Thus selfing rates are dependent on plant fitness.
Inbreeding can affect a plant’s mating system and this is called “the evolution of mating systems” and can be seen in the difference between monoecious and dioecious types.
Dioecious plants actually appear to be a development of monoecious plants.
The development of a dioecious plant is a two-step process. The initial step in the development of a dioecious plant is the “single gene male sterility mutation” which stops pollen production. The condition is known as “Gynodioecy” and the only reproductive parts that are functional in these monoecious plants are the female flowers. In this type of population there are female plants with sterile male sex organs and hermaphrodite plants. The male sterility gene will spread once a male sterility mutation combines with the population.
Cannabis in Androdioecy becoming male, cannabis in Gynodioecy becoming female and the original primordial hermaphrodite cannabis plant template.
“Androdioecy” is the related condition where a species has both male flowers with sterile
female
sex organs and hermaphrodite flowers, but this is a rare condition.
The next step in dioecious development is where a mutation of the pollen production at this locus occurs in the non-gynodioecious plants. The plants will try to stabilize this increase of males and in turn will reduce ovule production. As the modification takes place there is an equalized decrease in ovule production of the hermaphrodites over all the females. If hermaphrodites with increasing male function are selected, this gene will spread in the population. The resulting offspring will be females (with nonfunctional male organs) and males (with nonfunctional female organs). As time goes by the sex organs will evolve into either male or female sex organs and this is where the plants split into discrete sex types, which are dioecious. That is quite interesting isn’t it? It also goes some way to explaining why the sexual inheritance in cannabis and the expression of that sex is a complex subject.
Inbred lines have an adaptive value that can be seen in both the wild and in breeding labs. Inbreeding can result in populations that have higher levels of fitness because they are more suited to the environment that they have been inbreeding in.
Selfing has reproductive advantage and also resembles the inbred lines’ adaptive values. Although inbreeding results in a depression, that depression is only expressed if the inbred line’s environment is compromised or equilibrium has been altered. In order to be fit for such a problem a species will adapt through the chances of hybrid vigor when foreign genes enter the pool. We must also remember that inbreeding does not necessarily mean that deleterious genes will be fully eradicated. Homozygosity is increased and deleterious recessive alleles can be homozygous. To be realistic, the only time that deleterious genes will be removed is when these alleles are flushed from the population through the pressures of selection.