The Act of Creation (65 page)
Read The Act of Creation Online
Authors: Arthur Koestler

development; in the adult they are superseded by the integrative action
of the nervous system -- unless the embryonic potentials are reactivated
by regenerative needs. Similarly, the adult's mental co-ordination relies
on conscious, verbalized, 'logical' codes; not on the quasi 'embryonic'
(infantile, pre-causal) potentials of the unconscious; again unless these
are revived under the creative stress. Physical regenerations strike
us as 'spectacular pieces of magic' because they derive from prenatal
skills; and creative inspirations are equally mysterious because they
derive from levels which predate the conscious mind. As Polànyi
wrote (in a different context): 'The highest forms of originality are
far more closely akin to the lowest biotic performances than the external
circumstances would indicate.' [11]
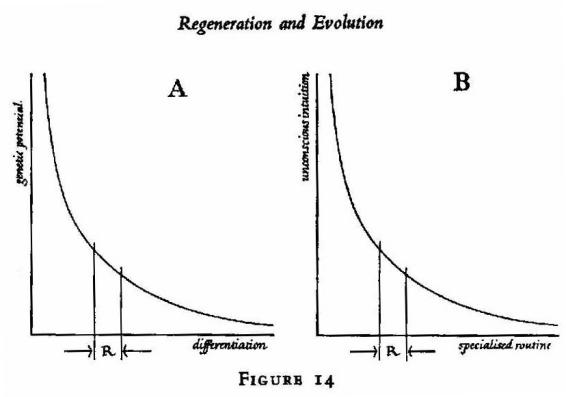
way the complementary factors in the
reculer pour mieux sauter
phenomenon. In A, increase in tissue-differentiation entails a reciprocal
decrease of genetic multipotentiality. In B, an analogous reciprocity
prevails between unconscious intuitions and automatized routines --
or, if you like, between fluid imagery and 'misplaced concreteness'. R
indicates the 'regenerative span'. (The curve in A should of course have
breaks and a series of discrete steps.)
It could be objected that structural regenerations merely restore
the
status quo ante
whereas mental reorganization leads to an
advance. But in the first place this is not always the case. Psychotherapy
aims at correcting 'faulty integrations' caused by traumatic experiences
-- at restoring normality. In the second place the biological evolution
of the species with which we are concerned has to all intents and
purposes come to a standstill, whereas mental evolution continues,
and its vehicle is precisely the creative individual. The Eureka
process is a mental mutation, perpetuated by social inheritance. Its
biological equivalent are the genetic mutations which carried the
existing species up the evolutionary ladder. Now a mutation -- whatever
its unknown cause -- is no doubt a re-moulding of previous structures,
based on a de-differentiation and reintegration of the otherwise rigid
genetic code. The transformations of fins into legs, legs into arms,
arms into wings, gills into lungs, scales into feathers, etc., while
preserving certain basic structural patterns (see, for instance, d'Arcy
Thompson's
On Growth and Form
) were eminently 'witty' answers
to the challenges of environment. It seems obvious that the dramatic
release, at periods of adaptative radiations, of unexplored morphogenetic
potentials by a re-shuffling of molecules in the genetic code, resulting
in the de-differentiation and reintegration of structures like limbs into
wings, is of the very essence of the evolutionary process. After all,
'ontogenesis and regenesis are components of a common mechanism', [12]
which must have a phylogenetic origin.
In Book One,
Chapter XX
, I have mentioned the
perennial myth of the prophet's and hero's temporary isolation and
retreat from human society -- followed by his triumphant return endowed
with new powers. Buddha and Mohamed go out into the desert; Joseph is
thrown into the well; Jesus is resurrected from the tomb. Jung's 'death
and rebirth' motif, Toynbee's 'withdrawal and return' reflect the same
archetypal motif. It seems that
reculer pour mieux sauter
is a
principle of universal validity in the evolution of species, cultures,
and individuals, guiding their progression by feedback from the past.
p. 454
. It is still an open question, however,
whether or how much undifferentiated 'reserve cells' (as in lower animals)
contribute material to the blastema.
p. 454
. It seems that the initial role of
the nervous system is to determine the main axis of the regenerate --
that it acts, not as an inductor, but as a trophic agent. At the later,
anabolic stages of the process no nerve supply is needed -- as denervation
experiments show.
p. 456
. This, actually, is the only clearly
demonstrated case of metaplasia among higher animals.
p. 459
. About the ways how this is achieved,
cf. McCulloch in the Hixon Symposium, p. 56.
represented by an ascending hierarchy of spatial levels perpendicular
to the time axis. In spite of the perplexing diversity of phenomena on
different levels -- cleavage, gastrulation, induction, neuro-genesis --
certain basic principles were seen to operate on every level throughout
the hierarchy. Principles (or 'laws of nature') can only be described
in symbolic language of one kind or another. The language used in the
present theory is based on four key-concepts: motivation, code, matrix,
and environment. Since these are assumed to operate in a hierarchic
framework, the dichotomy of self-asserting and participatory tendencies
of behaviour on all levels need not be separately postulated, but derives
logically, as it were, from the dual character of every sub-whole as a
sub-ordinate and supra-ordinate entity. Let me now recapitulate some of
the main points which have emerged from the previous chapters, taken in
conjunction with
Book One
:
J. Needham's tongue-in-the-cheek phrase about 'the striving of the
blastula to grow into a chicken' indicates the directiveness of the
morphogenetic process and its equifinal, regulative properties, which
become particularly evident under adverse conditions. These properties
represent the genetic precursors of the motivational drives, needs,
and goal-directedness of the adult animal; during maturation, the former
shade into the latter, and there is no sharp dividing line between them.
'whole'. Facing downward or outward in the hierarchy, it behaves as an
autonomous whole; facing upward or inward, it behaves as a dependent part
which is inhibited or triggered into action by higher controls. One might
call this the 'Janus principle' in organic (and social) hierarchies.
autonomous
and
spontaneous
activities. The principle of
autonomy is asserted on every level; from cell-organelles functioning as
power plants or motor units, through the self-differentiating activities
of the morphogenetic field, to the autonomous regulations of organs and
organ system. In the motor hierarchy, it is reflected at every stage, from
the muscle's selective response to specific excitation-patterns, through
the stubborn behaviour of the reversed newt limb, to the unalterable
features in a person's gait or handwriting. I have briefly mentioned (Book
One, Chapters
XIII
,
XXI
),
and shall discuss in more detail later, autonomous mechanism in perceptual
organization -- visual constancies, the automatic filtering, analysing,
generalizing of the input. Lastly, thinking and communicating are based
on hierarchically ordered, autonomous patterns of enunciation, grammar,
logic, mathematical operations, universes of discourse.
dynamic
aspect of the part's autonomy is manifested in
its apparently spontaneous, unprovoked rhythmic activities which are
'modified but not created by the environmental input' -- a statement
which equally applies to morphogenesis, to intrinsic motor patterns,
to the spontaneous discharges of unstimulated sensory receptors, the
electric pulses of the unstimulated brain, to drives in the absence of
external stimuli or to communications addressed to imaginary audiences.
'self-assertive' aspect of part-behaviour.
which may be said to represent the interests of the whole vis-à-vis
the part in question. The controls are largely of an inhibitory or
restraining character, to prevent overloading of information channels,
over-shooting of responses, confusion and redundancy in general; while
the activation of the part is effected by signals of the trigger-release
type. During morphogenesis, control is exercised by the suppression of
unrequired, and the release of required genetic potentials; in the mature
organism by interlacing multiple hierarchies of nervous and circulatory
processes, and biochemical gradients.
in supra-ordination to their own parts, (b) in sub-ordination to their
controlling agency, and (c) in co-ordination with their environment.
the environment of the nucleus is cytoplasm; on the level of the
morphogenetic field, the environment of one cell-population is another
cell-population; each organ in the adult animal is bathed in body-fluids
which constitute its environment. The structure and function of any
sub-whole is
determined
by (a) and (b) -- its intrinsic pattern
and its controls in the 'vertical' hierarchy to which it belongs; but it
is also
affected
by inputs and feedbacks from its 'horizontal'
environment, as it were -- the lie of the land. The difference between
(a) and (b) on the one hand, and (c) on the other is that the former
determine the invariant pattern of the operation, (c) only its variable
details. To mention a few examples: feedback from the cytoplasm to the
genetic blue-print co-determines into which variety of specialized cell
that particular unit will develop; but it does not alter the blue-print
itself. Inductor substances in the immediate environment of a tissue
will promote its differentiation into an organ -- but only within the
limits of the tissue's 'competence'. Environmental hazards decide the
neuro-muscular connections in the grafted salamander-limb, but its
functional co-ordination remains unaffected by it: it is controlled by
its 'vertical' hierarchy, and the lie of the land in the scar-tissue
determines only the local 'tactics' of the outgrowing nerve-filaments.
two types of 'input'. The first consists of specific trigger signals
from its superior controls in the hierarchy; the second are inputs
and feedbacks from more or less random events in its environment. But
I must stress again that the meaning of the word 'environment' depends
on the hierarchic level to which it is applied. The environment of John
driving his car is the traffic stream around him. The environment of
John's right foot is the brake-pedal on which it rests. Let us call
the former an environment on the t-level, where t stands for the top
of the hierarchy controlling the various sensory, motor, and cognitive
processes which constitute the skill of driving; then the brake-pedal
will be an environment on the, say, t-minus-4 level. Now John approaches
a sign which reads 'Halt -- Road Works Ahead'. This input is analysed and
relayed by various stations of the perceptual and cognitive hierarchy,
and is eventually re-coded into a 'sign releaser' which triggers off the
pre-set patterns of slow-down-to-a-halt behaviour on lower echelons of
the motor hierarchy. Thus an
environmental
input on the t-level has
been transformed into a specific trigger signal in a 'vertical' hierarchy;
in other words a 'category c' input on a higher level has been translated
into a 'category b' input on a lower level, activating the autonomous
pattern of the slowing-down skill. This consists in several sub-skills:
braking, steadying the wheel, going into neutral gear at the proper
moment. The foot on the brake-pedal is not responding to the 'Halt' signal
from the environment; it is responding to a specific 'excitation-clang'
travelling down John's spinal cord. But the foot's pre-set response is
modified by feedback from the environment on its own level: the 'feel'
of the pedal's elastic resistance governs the 'strategy' of braking --
neither too abruptly nor too softly. Similar feedbacks influence the
automatized motions of the hands on the wheel, etc.
Other books
Sea Mistress by Candace McCarthy
Wild Child: A Skull Kings MC Novella by Sage L. Morgan
The Remnants of Yesterday by Anthony M. Strong
The Winning Summer by Marsha Hubler
Parched by Georgia Clark
Rough Waters by Nikki Godwin
Modern Arrangements: Complete Trilogy (Modern Arrangements #1-3) by Sadie Grubor
Burn for You by Stephanie Reid
Buffalo Before Breakfast by Mary Pope Osborne
Impossible by Komal Lewis