Phantoms in the Brain: Probing the Mysteries of the Human Mind (14 page)
Read Phantoms in the Brain: Probing the Mysteries of the Human Mind Online
Authors: V. S. Ramachandran,Sandra Blakeslee
Tags: #Medical, #Neurology, #Neuroscience

If you turn the page upside down the eggs will transform themselves into cavities and cavities into eggs.
Adapted from Ramachandran, 1988a.
dinarily sophisticated processes in vision comes from neurology—from patients like Diane and others like her who have suffered highly selective visual deficits. If vision simply involves displaying an image on a neural screen, then in the case of neural damage, you would expect bits and pieces of the scene—or the whole scene—to be missing, depending on the extent of damage. But the defects are usually far more subtle than that. To understand what is really going on in the brains of these patients and why they suffer such peculiar 52

problems, we need to look more closely at the anatomical pathways concerned with vision.
Figure 4.3
The tail feathers of the argus pheasant have prominent disklike markings ordinarily shaded left to
right instead of top to bottom. Charles Darwin pointed out that when the bird goes through its courtship
ritual, the tail points up. The disks then are light on top
—
making them bulge out prominently like the eggs in
Figure 4.2. This may be the closest thing to the avian equivalent of jewelry.
From
The Descent of Man
by Charles Darwin (1871), John Murray, London.
When I was a student, I was taught that messages from my eyeballs go through the optic nerve to the visual cortex at the back of my brain (to an area called the primary visual cortex) and that this is where seeing takes place. There is a point−to−point map of the retina in this part of the brain—each point in space seen by the eye has a corresponding point in this map. This mapping process was originally deduced from the fact that when people sustain damage to the primary visual cortex—say, a bullet passes through one small area—they get a corresponding hole or blind spot in their visual field. Moreover, because of some quirk in our evolutionary history, each side of your brain sees the opposite half of the world (Figure 4.4). If you look straight ahead, the entire world on your left is mapped onto your right visual cortex and the world to the right of your center of gaze is mapped onto your left visual cortex.5
But the mere existence of this map does not explain seeing, for as I noted earlier, there is no little man inside watching what is displayed on
53
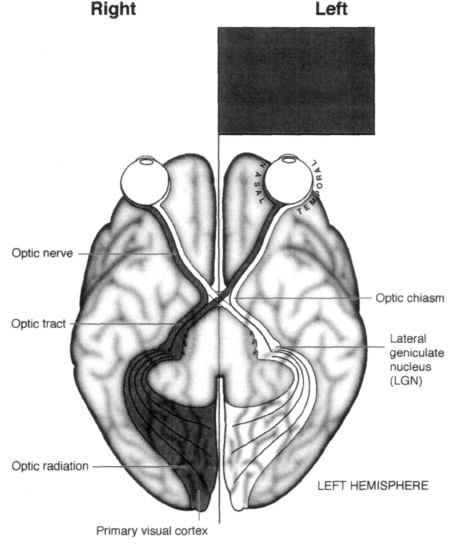
Figure 4.4
Bottom of the human brain viewed from below. Notice the curious arrangement of fibers going
from the retina to the visual cortex. A visual image in the left visual field (dark gray) falls on the right side of
the right eye's retina as well as the right side of the left eye's retina. The outer (temporal) fibers from the right
eye (dark gray) go then to the same right (visual) cortex without crossing at the optic chiasm. The inner
(nasal) fibers of the left eye (dark gray) cross at the chiasm and go to the right visual cortex as well. So the
right visual cortex "sees" the left side of the world.
Because there is a systematic map of the retina in the visual cortex, a "hole" in the visual cortex will cause a
corresponding blind spot (or scotoma) in the visual field. If the right visual cortex is completely removed, the
patient will be completely blind in the left side of the world.
Redrawn from S. Zeki,
A Vision of the Brain,
1993. Reproduced with permission from Blackwell (Oxford).
the primary visual cortex. Instead, this first map serves as a sorting and editorial office where redundant or useless information is discarded wholesale and certain defining attributes of the visual image—such as edges—are strongly emphasized. (This is why a cartoonist can convey such a vivid picture with just a few pen strokes depicting the outlines or edge alone; he's mimicking what your visual system is specialized to do.) This edited information is then relayed to an estimated thirty distinct visual areas in the human brain, each of which thus receives a complete or partial map of the visual world. (The phrases "sorting office" and "relay"
are not entirely appropriate since these early areas perform fairly sophisticated image analyses and contain massive feedback projections from higher visual areas. We'll take these up later.) 54
This raises an interesting question. Why do we need thirty areas?6 We really don't know the answer, but they appear to be highly specialized for extracting different attributes from the visual scene—color, depth, motion and the like. When one or more areas are selectively damaged, you are confronted with paradoxical mental states of the kind seen in a number of neurological patients. One of the most famous examples in neurology is the case of a Swiss woman (whom I shall call Ingrid) who suffered from "motion blindness." Ingrid had bilateral damage to an area of her brain called the middle temporal (MT) area. In most respects, her eyesight was normal; she could name shapes of objects, recognize people and read books with no trouble. But if she looked at a person running or a car moving on the highway, she saw a succession of static, strobelike snapshots instead of the smooth impression of continuous motion. She was terrified to cross the street because she couldn't estimate the velocity of oncoming cars, though she could identify the make, color and even the license plate of any vehicle. She said that talking to someone in person felt like talking on the phone because she couldn't see the changing facial expressions associated with normal conversation. Even pouring a cup of coffee was an ordeal because the liquid would inevitably overflow and spill onto the floor. She never knew when to slow down, changing the angle of the coffeepot, because she couldn't estimate how fast the liquid was rising in the cup. All of these abilities ordinarily seem so effortless to you and me that we take them for granted. It's only when something goes wrong, as when this motion area is damaged, that we begin to realize how sophisticated vision really is.
Another example involves color vision. When patients suffer bilateral damage to an area called V4, they become completely color−blind (this is different from the more common from of congenital color blindness that arises because color−sensitive pigments in the eye are deficient). In his book
An Anthropologist on Mars,
Oliver Sacks describes an artist who went home one evening after suffering a stroke so small he didn't notice it at the time. But when he walked into his house, all his color paintings suddenly looked as if they had been done in black and white. In fact, the whole world was black and white and soon he realized that the paintings had not changed, but rather something had happened to him. When he looked at his wife, her face was a muddy gray color—he claimed she looked like a rat.
So that covers two of the thirty areas—MT and V4—but what about all the rest? Undoubtedly they're doing something equally important, but we have no clear ideas yet of what their functions might be. Yet despite the bewildering complexity of all these areas, the visual system appears to have a relatively simple overall organization. Messages from the eyeballs go through the optic nerve and immediately bifurcate along two pathways—one phylogenetically old and a second, newer pathway that is most highly developed in primates, including humans. Moreover, there appears to be a clear division of labor between these two systems.
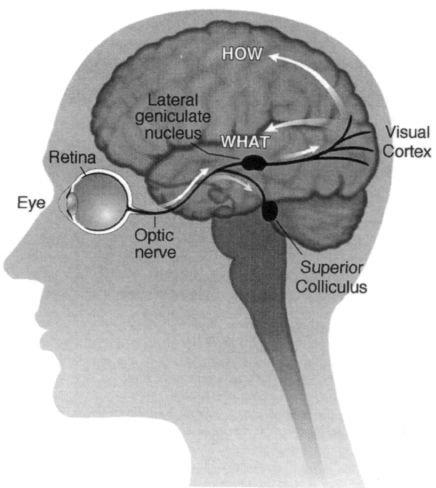
The "older" pathway goes from the eye straight down to a structure called the superior colliculus in the brain stem, and from there it eventually gets to higher cortical areas especially in the parietal lobes. The "newer"
pathway, on the other hand, travels from the eyes to a cluster of cells called the lateral geniculate nucleus, which is a relay station en route to the primary visual cortex (Figure 4.5). From there, visual information is transmitted to the thirty or so other visual areas for further processing.
Why do we have an old pathway and a new pathway?
One possibility is that the older pathway has been preserved as a sort of early warning system and is concerned with what is sometimes called "orienting behavior." For example, if a large looming object comes at me from the left, this older pathway tells me where the object is, enabling me to swivel my eyeballs and turn my head and body to look at it. This is a primitive reflex that brings potentially important events into my fovea, the high−acuity central region of my eyes.
At this stage I begin to deploy the phylogenetically newer system to determine what the object is, for only then can I decide how to respond to it. Should I grab it, dodge it, flee from it, eat it, fight it or make love to it?
55
Damage to this second pathway—particularly in the primary visual cortex—leads to blindness in the conventional sense. It is most commonly brought on by a stroke—a leakage or blood clot in one of the
Figure 4.5
The anatomical organization of the visual pathways. Schematic diagram of the left hemisphere
viewed from the left side.
The fibers from the eyeball diverge in two parallel "streams": a new pathway that goes to the lateral
geniculate nucleus (shown here on the surface for clarity, though it is actually inside the thalamus, not the
temporal lobe) and an old pathway that goes to the superior colliculus in the brain stem.
The "new" pathway then goes to the visual cortex and diverges again (after a couple of relays) into two
pathways (white arrows)
—
a "how" pathway in the parietal lobes that is concerned with grasping, navigation
and other spatial functions, and the second, "what" pathway in the temporal lobes concerned with recognizing
objects. These two pathways were discovered by Leslie Ungerleider and Mortimer Mischkin of the National
Institutes of Health. The two pathways are shown by white arrows.
main blood vessels supplying the brain. If the vessel happens to be a cerebral artery in the back of the brain, damage can occur in either the left or the right side of the primary visual cortex. When the right primary cortex is damaged, the person is blind in the left visual field, and if the left primary cortex is damaged, the right visual field is obliterated. This kind of blindness, called hemianopia, has been known about for a long time.
But it, too, holds surprises. Dr. Larry Weiskrantz, a scientist working at Oxford University in England, did a 56
very simple experiment that stunned experts on vision.7 His patient (known as D.B., whom I will call Drew) had an abnormal clump of blood vessels surgically removed from his brain along with some normal brain tissue in the same vicinity. Since the malformed clump was located in the right primary visual cortex, the procedure rendered Drew completely blind to the left half of the world. It did not matter whether he used his left eye or right eye—if he looked straight ahead, he could not see anything on the left side of the world. In other words, although he could see out of both eyes, neither eye could see its own left visual field.
After the surgery Drew's ophthalmologist, Dr. Mike Sanders, asked him to gaze straight ahead at a small fixation spot mounted in the center of a device that looks like an enormous translucent Ping−Pong ball.
Drew's entire visual field was filled with a homogeneous background. Next, Dr. Sanders flashed spots of light onto different parts of the curved screen mounted on the inside of a ball and asked Drew whether he could see them. Each time a spot fell into his good visual field, he'd say, "Yes, yes, yes," but when the spot fell into his blind region he would say nothing. He didn't see it.