Hen’s Teeth and Horse’s Toes (32 page)
Read Hen’s Teeth and Horse’s Toes Online
Authors: Stephen Jay Gould

But Bennett’s analysis is based upon only three characters, none very secure. All are potentially simple modifications of shape or proportion, not presences or absences of complex structures. All, like the sagittal crest, could come and go. Only one potential shared derived character unites true horses with
E. zebra:
the “orientation of postorbital bars relative to horizontal plane” (a relatively less slanted position for a bar of bone located on the skull behind the eyes—not exactly the stuff of which confident conclusions are made). Only two potential shared derived characters unite Burchell and Grevy zebras: the presence of frontal doming (inflation of the top part of the skull) and relative skull breadth (these two zebras have long and narrow snouts). Unfortunately, we know that at least one of these characters doesn’t work well for Bennett’s cladistic scheme because she admits that a member of her other lineage—a horse with the peculiar moniker of the Asiatic half-ass (
E. hemionus
)—has independently evolved a long, narrow snout. If twice, why not three times?
When we look for corroboration to an obvious source—numbers of chromosomes—we are again disappointed. As I discussed in essay 26, the various species of horses, despite their marked similarities of form, differ greatly in number of chromosomes. Fusion or fission of chromosomes may be a major mechanism of speciation in mammals, and these differences may therefore have great evolutionary significance. All zebras, and only zebras, have fewer than fifty pairs of chromosomes (thirty-two in Hartmann’s zebra to forty-six in Grevy’s zebra). All other horses have more than fifty (from fifty-six in
Equus hemionus
to sixty-six in Przewalski’s horse). The low number of zebras might mark them as a genealogical group if the character is shared derived, and not either primitive or evolved more than once. Bennett’s hypothesis may still be maintained by arguing either that small numbers are primitive for all horses and that asses and true horses acquired larger numbers by independent evolutionary routes; or that different lineages of zebras evolved small numbers along separate evolutionary paths. Still, since we have no reason to associate stripes with small numbers of chromosomes, their conjoined presence in all zebras might best be interpreted as a sign of genealogy. The more complex characters that a group shares, the more likely that the group is genealogical—unless we have good reason to regard all the characters as primitive (and we do not in this case).
I conclude that Bennett’s proposal is interesting, but very much unproven. Suppose, however, that she is right. What then would a zebra be? Or more specifically, how did cladistically unrelated horses get black-and-white stripes? There are two possibilities. Either the common ancestor of zebras and true horses had stripes and true horses lost them, while the three species of “zebras” passively retained them; or else, striping is an inherited developmental capacity of all horses and not so complex a character as it appears. In this case, several separate lineages could acquire stripes independently. Zebras would then be horses that have realized a potential pathway of development probably common to many or all members of the genus
Equus
(see next essay).
This particular tale of zebras may not hold, but the radical messages of cladistic ordering are secure in many cases. Some of our most common and comforting groups no longer exist if classifications must be based on cladograms. With apologies to Mr. Walton and to so many coastal compatriots in New England, I regret to report that there is surely no such thing as a fish. About 20,000 species of vertebrates have scales and fins and live in water, but they do not form a coherent cladistic group. Some—the lungfishes and the coelacanth in particular—are genealogically close to the creatures that crawled out on land to become amphibians, reptiles, birds, and mammals. In a cladistic ordering of trout, lungfish, and any bird or mammal, the lungfish must form a sister group with the sparrow or elephant, leaving the trout in its stream. The characters that form our vernacular concept of “fish” are all shared primitive and do not therefore specify cladistic groups.
At this point, many biologists rebel, and rightly I think. The cladogram of trout, lungfish, and elephant is undoubtedly true as an expression of branching order in time. But must classifications be based only on cladistic information? A coelacanth looks like a fish, tastes like a fish, acts like a fish, and therefore—in some legitimate sense beyond hidebound tradition
—is
a fish.
No debate in evolutionary biology has been more intense during the past decade than the challenge raised by cladistics against traditional schemes of classification. The problem arises from the complexity of the world, not from the fuzziness of human thought (although woolliness has made its usual contribution as well). We must recognize two rather different components to our vernacular conception of “similarity” between organisms—and classifications are designed to reflect relative degrees of similarity. On the one hand, we must consider genealogy, or branching order. Cladistics works with branching order alone, rigorously excluding any other notion of similarity. But what about the admittedly vague and qualitative, but not therefore unimportant, notion of overall similarity in form, function, or biological role? The coelacanth, to say it again, looks and acts like a fish even if its closer cladistic relatives are mammals. Another theory of classification, called phenetics—from a Greek word for appearance—focuses on overall similarity alone and tries to escape the charge of subjectivity by insisting that phenetic classifications be based upon large suites of characters, all expressed numerically and processed by computer.
Unfortunately, these two types of information—branching order and overall similarity—do not always yield congruent results. The cladist rejects overall similarity as a snare and delusion and works with branching order alone. The pheneticist attempts to work with overall similarity alone and tries to measure it in the vain pursuit of objectivity. The traditional systematist tries to balance both kinds of information but often falls into hopeless confusion because they really do conflict. Coelacanths are like mammals by branching order and like trout by biological role. Thus, cladists buy potential objectivity at the price of ignoring biologically important information. And traditionalists curry confusion and subjectivity by trying to balance two legitimate, but often disparate, sources of information. What is to be done?
I cannot answer this question, for it raises issues of style, mores, and methodology more than demonstrable substance. But I can at least comment on the source of this bitter debate—a rather simple point that somehow got lost in the heat. In an ideal world, there would be no conflict among the three schools—cladistics, phenetics, and traditional—and all would produce the same classification for a given set of organisms. In this pipe-dream world, we would find a perfect correlation between phenetic similarity and recency of common ancestry (branching order); that is, the longer ago two groups of organisms separated from a common ancestor, the more unlike they would now be in appearance and biological role. Cladists would establish an order of branching in time by cataloging shared derived characters. Pheneticists would crunch their numerous measures of similarity in their favorite computers and find the same order because the most dissimilar creatures would have the most ancient common ancestors. Traditionalists, finding complete congruence between their two sources of information, would join the chorused harmony of agreement.
But let the reverie halt. The world is much more interesting than ideal. Phenetic similarity often correlates very poorly with recency of common ancestry. Our ideal world requires a constancy of evolutionary rate in all lineages. But rates are enormously variable. Some lineages change not at all for tens of millions of years; others undergo marked alterations in a mere thousand. When the forebears of terrestrial vertebrates first split off from a common ancestry with coelacanths, they were still unambiguously fish in appearance. But they have evolved, along numerous lines during some 250 million years, into frogs, dinosaurs, flamingos, and rhinoceroses. Coelacanths, on the other hand, are still coelacanths. By branching order, the modern coelacanth may be closer to a rhino than a tuna. But while rhinos, on a rapidly evolving line, are now markedly different from that distant common ancestor, coelacanths still look and act like fish—and we might as well say so. Cladists will put them with rhinos, pheneticists with tunas; traditionalists will hone their rhetoric to defend a necessarily subjective decision.
Nature has imposed this conflict upon science by decreeing, through the workings of evolution, such unequal rates of change among lineages and such a poor correlation between phenetic similarity and recency of common ancestry. I do not believe that nature frustrates us by design, but I rejoice in her intransigence nonetheless.
SOME PERSISTENT
, unanswered questions about nature possess a kind of majestic intractability. Does the universe have a beginning? How far does it extend? Others refuse to go away because they excite a pedestrian curiosity but seem calculated, in their very formulation, to arouse argument rather than inspire resolution. As a prototype for the second category, I nominate: Is a zebra a white animal with black stripes or a black animal with white stripes? I once learned that the zebra’s white underbelly had decided the question in favor of black stripes on a blanched torso. But, to illustrate once again that “facts” cannot be divorced from cultural contexts, I discovered recently that most African peoples regard zebras as black animals with white stripes.
In a poem about monkeys, Marianne Moore discussed some compatriots at the zoo and contrasted elephants and their “strictly practical appendages” with zebras “supreme in their abnormality.” Yet we learned in essay 28 that the three species of zebras may not form a group of closest relatives—and that stripes either evolved more than once or represent an ancestral pattern in the progenitors of true horses and zebras. If stripes are not the markers of a few related oddballs but a basic pattern within a large group of animals, then the problems of their construction and meaning acquire more general interest. J. B. L. Bard, an embryologist from Edinburgh, has recently analyzed zebra stripes in the broad context of models for color in all mammals. He detected a developmental unity underlying the different patterns of adult striping among our three species of zebras and,
inter alia
, even proposed an answer to the great black-and-white issue in favor of the African viewpoint.
Biologists follow a number of intellectual styles. Some delight in diversity for its own sake and spend a lifetime describing intricate variations on common themes. Others strive to discover an underlying unity behind the differences that sort these few common themes into more than a million species. Among searchers for unity, the Scottish biologist and classical scholar D’Arcy Wentworth Thompson (1860–1948) occupies a special place. D’Arcy Thompson spent his life outside the mainstream, pursuing his own brand of Platonism and packing insights into his thousand-page classic,
On Growth and Form—
a book so broad in appeal that it won him an honorary degree at Oxford and, thirty years later, entered the
Whole Earth Catalog
as “a paradigm classic.”
D’Arcy Thompson struggled to reduce diverse expressions to common generating patterns. He believed that the basic patterns themselves had a kind of Platonic immutability as ideal designs, and that the shapes of organisms could only include a set of constrained variations upon the basic patterns. He developed a theory of “transformed coordinates” to depict variations as expressions of a single pattern, stretched and distorted in various ways. But he worked before computing machines could express such transformations in numerical terms, and his theory achieved little impact because it never progressed much beyond the production of pretty pictures.
As a subtle thinker, D’Arcy Thompson understood that emphases on diversity and unity do not represent different theories of biology, but different aesthetic styles that profoundly influence the practice of science. No student of diversity denies that common generating patterns exist, and no searcher for unity fails to appreciate the uniqueness of particular expressions. But allegiance to one or the other style dictates, often subtly, how biologists view organisms and what they choose to study. We must reverse the maxim of the reprobate father teaching his son morality—do what I say, not what I do—and recognize that biological allegiances lie not so much in words but in actions and subjects chosen for research. Note what I do, not what I say. Of the “pure taxonomist”—the describer of diversity—D’Arcy Thompson wrote:
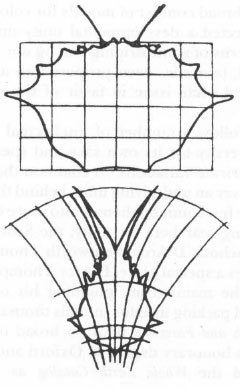
“Transformed coordinates” on the carapaces of crabs from two different genera demonstrate unity of form.
FROM ON GROWTH AND FORM, BY D’ARCY WENTWORTH THOMPSON/CAMBRIDGE UNIVERSITY PRESS
, 1917.
When comparing one organism with another, he describes the differences between them point by point and “character” by “character.” If he is from time to time constrained to admit the existence of “correlation” between characters…he recognizes this fact of correlation somewhat vaguely, as a phenomenon due to causes which, except in rare instances, he can hardly hope to trace; and he falls readily into the habit of thinking and talking of evolution as though it had proceeded on the lines of his own description, point by point and character by character.
D’Arcy Thompson recognized, with sadness, that the theme of underlying unity had received much lip service, but little application. Differences between the striping patterns of our three zebra species had been described minutely and much energy had been invested in speculations about the adaptive significance of differences. But few had asked whether all the patterns might be reduced to a single system of generating forces. And few seemed to sense what significance such a proof of unity might possess for the science of organic form.
The vulgar version of Darwinism (not Darwin’s) holds that natural selection is so powerful and pervasive in scrutinizing every variation and constructing optimal designs that organisms become collections of perfect parts, each minutely crafted for its special role. While not denying correlation in development or underlying unity in design, the vulgar Darwinian does relegate these concepts to unimportance because natural selection can always break a correlation or remold an inherited design.
The pure vulgar Darwinian may be a fiction; no one could be quite so foolish. But evolutionary biologists have often slipped into the practice of vulgar Darwinism (while denying its precepts) by following the reductionistic research strategy of analyzing organisms part by part and invoking natural selection as a preferred explanation for all forms and functions—the point of D’Arcy Thompson’s profound statement cited above. Only in this way can I make sense of the curious fact that unity of design has received so little attention in the practice of research—although much lip service in textbooks—during the past forty years, while evolutionary biologists have generally preferred a rather strict construction of Darwinism in their explanations of nature.
For many reasons, ranging from the probable neutrality of much genetic variation to the nonadaptive nature of many evolutionary trends, this strict construction is breaking down, and themes of unity are receiving renewed attention. Old ideas are being rediscovered; D’Arcy Thompson, although never out of print, is now often out of bookstores (and in personal libraries). One old and promising theme emphasizes the correlated effects of changes in the timing of events in embryonic development. A small change in timing, perhaps the result of a minor genetic modification, may have profound effects on a suite of adult characters if the change occurs early in embryology and its effects accumulate thereafter.
The theory of human neoteny, often discussed in my essays (see my disquisition on Mickey Mouse in
The Panda’s Thumb
) is an expression of this theme. It holds that a slowdown in maturation and rates of development has led to the expression in adult humans of many features generally found in embryos or juvenile stages of other primates. Not all these features need be viewed as direct adaptations built by natural selection. Many, like the “embryonic” distribution of body hair on heads, armpits, and pubic regions, or the preservation of an embryonic membrane, the hymen, through puberty, may be nonadaptive consequences of a basic neoteny that is adaptive for other reasons—the value of slow maturation in a learning animal, for example.
Bard’s proposal for “a unity underlying the different zebra striping patterns” follows D’Arcy Thompson’s theme of a basic motif stretched and pulled in different ways by varying forces of embryonic growth. These varying forces arise because the basic pattern develops at
different times
in the embryology of the three species. Bard thus combines the theme of transformed coordinates with the insight that substantial evolution can proceed by changes in the timing of development.
The basic pattern is simplicity itself: a series of parallel stripes deposited perpendicular to a line running along the embryonic zebra’s back from head to tail—hang a sheet over a taut wire and paint vertical stripes on each side of it. These stripes are initially laid down at a constant size, no matter how big the embryo that forms them. They are 0.4 mm., or approximately 20 cell diameters, apart. The bigger the embryo, the greater the initial number of stripes. (I should point out that Bard’s argument is a provocative model for testing, not a set of observations; no one has ever traced the embryology of zebra striping directly.)
The three zebra species differ in both number and configuration of stripes. In Bard’s hypothesis, these complex variations arise only because the same basic pattern—the parallel stripes of constant spacing—develops during the fifth week of embryonic growth in one species, during the fourth week in another, and during the third week in the third species. Since the embryo undergoes complex changes in form during these weeks, the basic pattern is stretched and distorted in varying ways, leading to all the major differences in adult striping.
The three species differ most notably in patterns of striping on the rump and hind quarters (see illustration in the last chapter). Grevy’s zebra
(Equus grevyi)
has numerous fine and basically parallel stripes in these rear regions. On Bard’s model, the stripes must have formed when the back part of the embryo was relatively large. (The larger the part, the more stripes it receives, since stripes are initially formed at constant size and spacing.) In the embryology of horses, the tail and hind regions expand markedly during the fifth week
in utero
. If adults possess numerous, fine posterior stripes, they must form after this embryonic expansion of the rear quarters. (Unfortunately, no one has ever studied the early embryology of zebras directly, and Bard assumes that the intrauterine growth patterns of true horses are followed by their striped relatives as well. Since basic features of early embryology tend to be highly conservative in evolution, true horses are probably fair models for zebras.)
The mountain zebra,
Equus zebra
, looks much like
E. grevyi
until we reach the haunch, where three broad stripes substitute for the numerous fine stripes of Grevy’s zebra. Broad stripes on adults indicate initial formation on a small piece of embryo (where few stripes could fit), and later rapid growth of the piece (widening the stripes as the general area expands). If an embryo forms stripes in its fourth week, just before the posterior expansion that provides room for the many fine marks of Grevy’s zebra, it will build the pattern of a mountain zebra during later embryonic growth.
Burchell’s zebra,
Equus burchelli
, also has just a few broad stripes on its haunch. But, while the mountain zebra has fine stripes over most of its back and broad stripes only over the haunch, the broad stripes of Burchell’s zebra begin in the middle of the belly and sweep back over the haunch. This pattern suggests an initial formation of stripes during the third week of embryonic growth. At this early stage, the embryo has a short, compact back, which later expands toward the rear in a broad, arching curve while the belly remains short. A stripe that initially ran vertically from belly to spine would be pulled toward the rear as the embryo’s top surface expanded backward while its belly grew little. An adult stripe, subject to such deformation in its embryonic life, would be broad and would run from the belly up and over the haunch—as in Burchell’s zebra.
Thus, Bard can render differences in rear striping of all three species as the results of deforming the same initial pattern at different times during normal embryonic growth. His hypothesis receives striking support from another source: the total number of stripes itself. Remember that Bard assumes a common size and spacing for stripes at their initial formation. Thus, the larger the embryo when stripes first form, the greater the number of stripes. Grevy’s zebra, presumably forming its stripes as an embryo of five weeks and about 32 mm. in length, has eighty or so stripes as an adult—or about 0.4 mm. per stripe. Mountain zebras with a fourth-week embryo of some 14 to 19 mm. have about forty-three stripes—again about 0.4 mm. per stripe. Burchell’s zebra has twenty-five to thirty stripes; if they form in a third-week embryo some 11 mm. long, we get the same value—about 0.4 mm. per stripe.
As additional support, and a lovely example of the difference between superficial appearance and knowledge of underlying causes, consider an old paradox involving hybrid offspring between zebras and true horses. These animals almost always have more stripes than their zebra parent. “Common sense,” based on superficial appearance, declares this result puzzling. After all, the state between stripes and no stripes is few stripes. But if Bard is right about the underlying causes of striping, then this paradoxical result makes sense. The intermediate state between stripes and no stripes might well be a
delay
in the embryonic formation of stripes. If stripes then begin at their common size and spacing upon a larger embryo, the resulting adult will have
more
stripes.