Connectome (14 page)
Authors: Sebastian Seung

Â
 Â
Â
Â
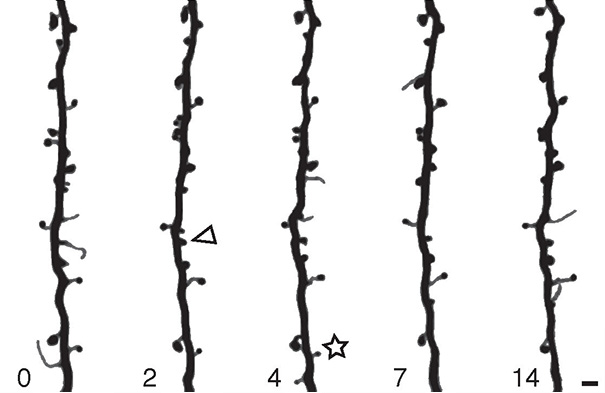
Figure 23. Evidence for reconnection: spines appearing and disappearing on a dendrite in the cortex of a mouse
Â
The dendrite bears thornlike protuberances known as spines. Most synapses between excitatory neurons are made onto spines rather than onto the shaft of the dendrite. In the figure, some spines are stable for the whole two weeks, but others appear (for example, look at the spine indicated by the arrowhead) and disappear (see the starred spine). This is good evidence that synapses are being created and eliminated.
Researchers still debate how frequently such reconnection happens, but all agree that it is possible.
Why are reweighting and reconnection so important? These two types of connectome change continue to happen for our entire lives. We must study them if we want to understand personal change as a lifelong phenomenon. No matter how old we get, we never stop storing new memories, barring some kind of brain disorder. As we age, we may complain that it's more difficult to learn, but even the elderly can acquire new skills. It seems likely that reweighting and reconnection are involved in such changes.
But do we have any proof? Evidence implicating reweighting in memory storage has come from Eric Kandel and his collaborators, who studied the nervous system of
Aplysia californica,
a squishy creature found in tide pools of California beaches. This animal retracts its gill and siphon when disturbed, and can become more or less sensitive to disturbancesâa simple kind of memory. We previously learned that such behaviors depend on neural pathways from sense organs to muscles. Kandel identified a single connection in the relevant pathway and showed that changes in its strength were related to the simple memory mentioned above.
Is reconnection involved in memory storage? Earlier I mentioned the phrenological idea of learning as thickening of the cortex. In the 1970s and 1980s, William Greenough and other researchers found evidence that such thickening was caused by an increase in the number of synapses. Their findingsâwhich were made by counting synapses
in the thickened cortex of rats who had been raised in enriched cagesâled some to propose a neo-phrenological theory:
Memories are stored by creating synapses.
Neither of these approaches truly succeeded in elucidating memory storage, however. Kandel's approach has faltered for brains more like our own, in which memories do not appear localized to single synapses. It seems more probable that memories are stored as patterns of many connections. Greenough's approach is also incomplete, because counting synapses does not tell us how they are organized into patterns. Furthermore, increases in synapse number, like cortical thickening, are correlated with learning, but it's not clear whether they are causally related.
To really crack the problem of memory, we need to figure out whether reweighting and reconnection are involved, and if so, exactly how. Earlier I explained the theory that the patterns of connection relevant for memory are cell assemblies and synaptic chains. Here I'll take a further step and propose that these patterns are created by reweighting and reconnection, and I'll explore the many questions that arise. Are these two processes independent, or do they work together? Why would the brain use both rather than just one? Can we explain some limitations of memory as malfunctions of these storage processes?
Beyond satisfying our basic curiosity about memory, research on reweighting and reconnection could have practical consequences. Suppose that your goal is to develop a drug that improves memory storage. If you believe neo-phrenology, you might try to develop a drug that enhances the molecular processes involved in synapse creation. But if neo-phrenology is wrongâas it most likely isâyour creation of more synapses might have effects very different from what you intended. More generally, whether we want to improve our memory abilities or prevent them from malfunctioning, knowledge about the basic mechanisms will be essential.
Â
We've seen how a cell assembly might retain associations between ideas as connections between neurons. But how does the brain create a cell assembly in the first place? This is the connectionist version of a much older question posed by philosophers: Where do ideas and their associations come from? While some might be innate, it's clear that others must be learned from experience.
Over the ages, philosophers came up with a list of principles by which associations can be learned. At the top of the list is coincidence, sometimes called contiguity in time or place. If you see photos of a pop singer with her baseball-player boyfriend, you will learn an association between them. A second factor is repetition. Seeing these celebrities together just once might not be enough to create the association in your mind, but if you see them ad nauseam day after day in every magazine and newspaper, you will not be able to avoid learning the association. Ordering in time also seems important for some associations. As a child you recited the letters of the alphabet repeatedly until you knew them by heart. You learned the association from each letter to the next, since the letters always followed one another in the same sequence. In contrast, the association between the pop singer and her boyfriend will be bidirectional, since they always appear simultaneously.
So philosophers proposed that we learn to associate ideas when one repeatedly accompanies or succeeds another. This inspired connectionists to conjecture:
Â
If two neurons are repeatedly activated
simultaneously,
then the connections between them are strengthened in both directions.
This rule of plasticity is appropriate for learning two ideas that repeatedly occur together, like the pop singer and her boyfriend. For learning associations between sequential ideas, connectionists proposed a similar rule:
Â
If two neurons are repeatedly activated
sequentially,
the connection from the first to the second is strengthened.
In both rules, by the way, it's assumed that the strengthening is permanent or at least long-lasting, so that the association can be retained in memory.
The sequential version of the rule was hypothesized by Donald Hebb,
who also proposed the cell assembly in his 1949 book,
The Organization of Behavior.
Both simultaneous and sequential versions have come to be known as Hebbian rules of synaptic plasticity. Both are said to be “activity-dependent,” because plasticity is triggered by the activity of the neurons involved in the synapse. (There are other ways of inducing synaptic plasticity that do not involve activity, such as the application of certain drugs.) Typically, Hebbian plasticity refers
only to synapses between excitatory neurons.
Hebb was way ahead of his time. Neuroscientists had no means of detecting synaptic plasticity. In fact, they could not even measure synaptic strengths at all. Measurements of spiking had been conducted for decades using metal wires inserted into the nervous system. Since the tip of the wire remained outside the neuron, this method was known as “extracellular” recording. The signals from the wire carried the spikes of several neurons, mixing them together like conversations in a crowded bar. This method, still in use today, is the one that was employed by Itzhak Fried and his collaborators to find the “Jennifer Aniston neuron.” By carefully maneuvering the tip of the wire, it's possible to isolate the spikes
of a single neuron, much as you do when you stick your ear close to the mouth of one of your friends at the bar.
While extracellular recording was sufficient for detecting spikes, it failed to measure the weak electrical effects of individual synapses. This was first accomplished in the 1950s by inserting a glass electrode with an extremely sharp tip into a single neuron. Such “intracellular” recording is so precise that it can detect signals much weaker than spikes, the equivalent of sticking your ear
inside
the mouth of a speaker
at a bar. An intracellular electrode can also be used to stimulate a neuron to spike, by injecting electrical current
into the neuron.
To measure the strength of a synapse from neuron A to neuron B, we insert electrodes into both neurons; we stimulate neuron A to spike, which causes the synapse to secrete neurotransmitter; and we measure the voltage of neuron B, which responds with a blip. The size of this blip
is the strength of the synapse.
Along with measuring a synapse's strength, we can also measure
changes
in its strength. To induce Hebbian plasticity, we stimulate spiking in a pair of neurons. Repeated stimulation,
either sequential or simultaneous, has been shown to strengthen synapses in accordance with the two versions of the Hebbian rule given earlier.
After a change in synaptic strength has been induced, it can last for the rest of the experimentâa few hours at most, as it's not easy to keep the neurons alive after they've been penetrated with electrodes. But cruder experiments involving populations of neurons and synapses, first done in the 1970s, suggest that changes in synaptic strength can last for weeks or longer. The issue of persistence is critical if Hebbian plasticity is to be the mechanism of memory storage, as some memories can last for a lifetime.
These experiments from the 1970s provided the first evidence for synaptic strengthening. By that time a theory of memory storage had also emerged, based on Hebb's original ideas. In the simplest version of the theory, a network starts out with weak synapses in both directions between the neurons of every pair. This assumption will turn out to be problematic, but let's accept it for now, for the purpose of introducing the theory.
Return to the scene of your first kiss, the actual event that imprinted your memory. The “magnolia neuron,” the “red brick house neuron,” the “sweetheart neuron,” the “plane neuron,” and so on were being activated by the stimuli around youâquite vigorously, I imagine. If we assume the simultaneous version of the Hebbian rule, all this spiking strengthened the synapses between these neurons.
The strengthened synapses together constitute a cell assembly, if we redefine this concept to mean a set of excitatory neurons mutually interconnected by
strong
synapses. Our original definition didn't have this stipulation. We need it now because the network contains many weak synapses that do not belong to the cell assembly. They existed before your first kiss, and remained unchanged afterward.
The weak synapses have no effect on recollection. Activity spreads from neuron to neuron within the cell assembly but does not spread any farther, because synapses from the cell assembly to other neurons are too weak to activate them. Thus the new definition of a cell assembly functions just as the old one did.
An analogous theory applies for the synaptic chain. Suppose that a sequence of stimuli activates a sequence of ideas. Each idea is represented by the spiking of a group of neurons. If the groups spike in this sequence repeatedly, the sequential version of the Hebbian rule will strengthen all existing synapses from neurons in each group to neurons in the next group. This is a synaptic chain, if we redefine this concept to mean a pattern of
strong
connections.
If the connections are sufficiently strong, then the spiking will propagate through the chain without any need for a sequence of external stimuli. Any stimulus that activates the first group of neurons will trigger the recollection of a sequence of ideas, as described in Chapter 4. Every successive recollection of the sequence will further strengthen the connections of the chain by Hebbian plasticity. This is analogous to the way that the flowing water of a stream slowly deepens its bed, making it even easier for the water to flow.
While it's important to remember things, it's also vital to forget. At one time your Jennifer Aniston and Brad Pitt neurons were linked by strong synapses into a cell assembly. But one day you started to see Brad with Angelina. (I know it was sad, but I hope you didn't feel
too
devastated.) Hebbian plasticity strengthened the connections between your Brad and Angelina neurons, creating a new cell assembly. What happened to the connections between your Brad and Jen
neurons?
You could imagine an analogue of the Hebbian rule serving the function of forgetting. Perhaps the connections between two neurons are weakened
if one is repeatedly active while the other is inactive. This would weaken the synapses between Brad and Jen every time you saw him without her.
Alternatively, one can imagine that weakening is caused by direct competition
between synapses. Perhaps the synapses between Brad and Angelina directly compete with those between Brad and Jen for some foodlike substance that synapses need in order to survive. If some synapses strengthen, they consume more of the substance, leaving less for the others, which grow weak. It's not clear whether such substances exist for synapses, but analogous “trophic factors”
are known to exist for neurons. Nerve growth factor is one example; its discovery won Rita Levi-Montalcini and Stanley Cohen a 1986 Nobel Prize.