A New History of Life (19 page)
Read A New History of Life Online
Authors: Peter Ward

One of the iconic fossils, trilobites were arthropods that dominated oceanic habitats relatively early in the history of animals on Earth. But how early? In Darwin’s time, trilobites were thought to be the earliest of all animals. Yet they are undoubtedly complicated, with three body sections, complex eyes and limbs—and large size. Some of the earliest could be up to two feet long. This was not what the earliest animals
ought
to have looked like—small and generalized, not large, complicated kinds of animals. We now know that trilobites were not—not even close, in fact, to being—the first animals.
3
The history of the origin of animals on Earth is one of life’s most fascinating chapters, and also one of the most controversial. There is also a great deal of new information that has been gleaned even in the last ten years. There are two distinct lines of evidence giving quite different views on the timing of the first diversification of animal phyla. One of these lines comes from the pattern of appearance of animal fossils in rocks, the second from molecular clock studies on extant animals. They give important clues to one of the greatest of all paleontological mysteries: the rapid diversification of animals.
Illustration of trilobites from the nineteenth century. At that time, these were thought to be the oldest fossils on Earth. Trilobites were used to “mark” the start of the Cambrian Period.
The first major line of evidence about the Cambrian explosion comes from fossils. The appearance of animals leaving evidence of themselves in the rock record came in four successive waves. The first began around 575 million years ago and has been called the Avalon explosion, a name coming from the part of eastern Canada where the oldest of this group were found. The second wave is coincident with the almost complete disappearance of the Ediacarans, and is characterized not by actual fossils but with accurate traces of their locomotion. These numerous “trace fossils” could only have been formed by active locomotion of multicellular organisms—animals. These are as old as 560 million years, but most are about 550 million years in age. The sea bottoms would have been alive with actively moving, small wormlike forms.
4
The third breakthrough was the appearance of skeletons, great numbers of tiny skeletal elements, in strata less than 550 million years in age. They are very small spines and scales of calcium carbonate that would have covered the animals with a coating of these small skeletons, almost like tiles. Finally, the larger fossilized animals appeared, including trilobites, the clam-like brachiopods, spiny echinoderms, and many kinds of snail-like mollusks, all in strata younger than 530 million years in age. In Darwin’s day, none of the earlier three were known, and the Cambrian was marked by the first appearance of trilobites in sedimentary strata. The reasons for this sequence might be deceptively simple: oxygen levels, which rose to their highest levels of the world up until then.
Today we know that this succession of animal life originations appeared comparatively rapidly in the fossil record, and new dating techniques now puts the time of the first complex
fossils
(the small skeletal fossils, which are 20 to 10 million years younger than the first trace fossils) at slightly older than 540 million years ago, with the first trilobites appearing in the record some 20 million years after that.
The appearance of animals in the fossil record recorded a significant event, which has been called the Cambrian explosion. To paleontologists the Cambrian explosion marked the first appearance
of most major animal phyla large enough to leave remains in the rock record. To molecular geneticists, the Cambrian marked the first evolution of animals. The controversy raged through the 1990s, to be solved in the early years of this century when new molecular studies
5
using more sophisticated analyses essentially confirmed the younger date for the origin of animals that had been championed by paleontologists. There is now agreement that animal life on Earth did not predate 635 million years ago,
6
and might be closer to 550 million years in age.
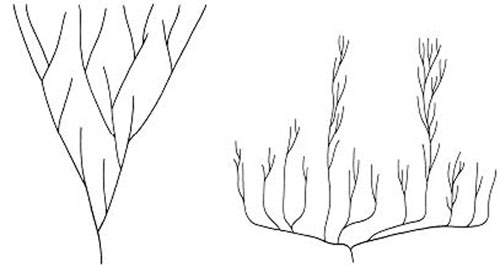
Left, the cone shape represents the traditional model for increasing disparity. Right, the inverted cone represents diversification and decimation.
The Cambrian period is now dated from 542 to about 495 million years ago (although the latter date, for the base of the Ordovician, might be slightly older). However, the vast majority of animal phyla first appeared in a small portion of this interval, between 530 and 520 MA. All specialists agree that this is the third or fourth most important event in the entire history of life, superseded in importance only by the first appearance of life on Earth, the adaptation to molecular oxygen, and the origin of the eukaryotic cell.
7
According to our best new information, the oxygen level soon after the start of the Cambrian explosion was about 13 percent (compared to 21 percent today),
8
but then fluctuated. During this time carbon dioxide levels were far higher than they are in the world
today—hundreds of times higher, in fact, and such high levels would have produced an intense greenhouse effect, sufficiently high to overcome the fact that the sun at this time was ~5 percent less intense than it is today. Even with the drop in CO
2
levels at the end of this interval, temperatures of this time would have been perhaps the highest of any period in the history of animal life on Earth. Since less oxygen is dissolved in seawater with higher temperatures, the already anoxic conditions of the oceans would have been exacerbated.
The panoply of fossils that have been preserved showing both hard and their soft-parts fossils from the fantastic and newly discovered deposits in the Chengjiang region of China has given us a new window into the origin of the animal phyla on Earth, and the nature of life on the Cambrian planet prior to the most famous of all fossil deposits, the Burgess Shale of British Columbia. The Chengjiang beds are now known to have been deposited between 520 and 515 million years ago, whereas the Burgess Shale is now thought to be no older than 505 million years in age. The approximately 10 million years separating the age of these two deposits thus gives us a new view of how animals diversified.
Because both Chengjiang and the Burgess preserve soft parts as well as skeletonized animals,
9
we have a good picture of what was there, in what relative abundance. Without this added view yielded by the preservation of soft parts, we would never be sure about the relative abundance of various kinds of animals, for perhaps there was a huge abundance of creatures like soft worms and jellyfish, forms that did not have skeletons. Thus our surprise at what appears to be a clear view of the nature of the fauna at both sites. There have now been over fifty thousand fossils collected from the Burgess Shale (and a lesser number of from Chengjiang). In their masterful summary of the Burgess fauna, Derek Briggs, Doug Erwin, and Fred Collier (in their 1994 book
The Fossils of the Burgess Shale
10
) list a total of 150 species of animals. Almost half are arthropods or arthropod-like. But an even more interesting number relates to the number of individuals. Well over 90 percent of all fossils are from arthropods, followed by sponges and brachiopods. Like the earlier Chengjiang, the Burgess
sea bottom was dominated both in kinds and numbers of animals by the arthropods.
Arthropods are among the most complex of all invertebrates, and yet, in these almost earliest of fossil deposits in the time of animals, they are diversified and common. It speaks to a long evolution prior to their first appearance in the record—perhaps seabeds crawling with millimeter-long (or less) arthropods, with many more species swimming or floating in the open sea itself.
One of the great surprises of a visit to the Burgess Shale (which both of the authors of this book have been fortunate enough to do) is the realization that the most common fossils come not from the exotic taxa, the many exquisite, soft-bodied creatures that fill the pages of the many books devoted to the Burgess Shale fauna and flora, but the fact that most of the fossils come from trilobites. They, and the less numerous but highly diverse arthropods of the Burgess dominate the assemblage,
11
in sheer numbers of individuals and species, and in sheer numbers of different kinds of body plans, which is described by a measure called disparity (and compared to diversity, which refers to the number of different kinds of taxa). The arthropods seem to have been the most successful of Cambrian animals. How much of this success was due to their principal body plan characteristic: segmentation?
Segmented animals are the most diverse of all animals on the planet, and most are arthropods. All arthropods, including the highly diverse insects, show repeated body units and body regions based on groupings of individual segments that have specific functions for the animal. The feature uniting the group is the presence of a jointed exoskeleton that encloses the entire body. This exoskeleton even extends into the gut. The exoskeleton cannot grow, so it must be periodically molted and replaced by another slightly larger one. The body has a well-differentiated head, trunk, and posterior regions in varying proportions. Appendages are commonly specialized. On terrestrial arthropods the appendages are usually single (enormous), but the marine forms generally have two branches or parts per appendage, an inner leg branch and an outer gill branch, and are thus termed
biramous. The exoskeleton encloses the soft parts like a suit of armor, and that may be its major function: protection. But the consequences of this kind of skeleton are huge: there can be no passive diffusion of oxygen across any part of the body. To obtain oxygen the first arthropods, all marine, had to evolve specialized respiratory structures or gills. Segmented animals are the most diverse of all animals on the planet. Arthropods are not alone in this trait: all annelids are segmented, and some members of generally nonsegmented groups, such as the monoplacophoran mollusks, show at least some segmentation. It appeared early in the history of animals, and indeed in the Cambrian trilobites we see that the most common of these early preserved animal fossils show this trait.
In his 2004 book,
On the Origin of Phyla
,
12
James Valentine also reflects on what is a major evolutionary puzzle: why are there so many, and so many kinds of arthropods in the Cambrian? It is worthwhile to look at what he has written on this subject:
Although many early arthropods had non-mineralized cuticles, a marvelous diversity of early arthropod body types has come to light, so many and so distinctive as to pose important problems in applying the principles of systematics. These disparate arthropod types are phylogenetically puzzling … This evidently sudden burst of evolution of arthropod-like body types is outstanding even among the Cambrian Explosion taxa.
What we call arthropods are composed of what appear to be perhaps many independently evolved groups that have, through convergent evolution, produced body plans of great diversity save for one aspect: all have limbs on each segment that are biramous—each appendage carries a leg of some sort, and a second appendage, a long gill.
Why would basal animal groups opt for segmentation? Perhaps this is the wrong word, for Valentine and others note that the arthropods are not so much segmented—which at least in annelids is composed of largely separated chambers for each segment of the
body—but repeated. Valentine proposes that this striking body plan arose in response to locomotor needs, stating, “Cleary, the segmented nature of the arthropod’s body is related to the mechanics of body movement, particularly to locomotion, with nerve and blood supplies in support.” There is no doubt that this type of body plan is an adaptation aiding locomotion. But a consequence of this kind of body plan is to allow repeated gill segments, each small enough to be held in optimal orientation beneath the segments. In these positions, flows of water can be actively pumped over and through the feather-shaped gills, thereby increasing the availability of oxygen molecules hitting the gills each second, a position suggested by Ward in 2006.
13